


With breathtaking panache, ostensibly natural, and comparable to unsullied oceanic reefs, aquaria dominated by SPS corals are ultrapure with intense irradiance and water movement that expedites growth.

Fig 1. Demersal green clown goby (Gobiodon histrio) alight on branching SPS colonies and remain small.
These corals require enduring stability and year-round temperatures of ~23.5oC, currents equal to the turnover of the entire display volume 70 times per hour, no more than a weekly 7 percent water change, intense photosynthetically useful radiation (PUR) with profound backscatter, nominally undetectable but post-feeding pulses of NH4+/NH3 and NO2-, nil sources of H2S and derivatives, low dissolved CO2 but ~11 degrees carbonate hardness (dKH), nominal planktonic bacteria, optimized dissolved oxygen (DO), pH, and oxidation-reduction potential (ORP), and ~0.75 and 0.065 mg l-1 NO3- and PO43- respectively. That said, such constraints are challenging to sustain.
These corals deposit calcium carbonate at an alarming rate with the correct alkalinity, illumination, and flow, and thus require sufficient depth because their cultivation is forestalled once they reach the surface. They can be kept in a fish-free system but they require ambient ammonia which they may gain from their zooplanktonic feeds that satisfy around 35 percent of their requirements (Houlbrèque & Ferrier‐Pagès 2009). Yet such administrations pollute and cause mortality without established communities of nitrogen cycling microorganisms.
Ammonia produced by small fish with quick metabolisms stabilizes nitrifying and denitrifying prokaryotic populations and thus water quality. It is wise therefore to house SPS colonies in aquaria at least three months old with a nominal but long-standing fish biomass of no more than 1” (25 cm) of fish to every 10 UK gallons. Green clown goby (Gobiodon histrio) reach merely 3 cm (1.2”) which makes them perfect for pico-SPS reefs (Fig 1.).
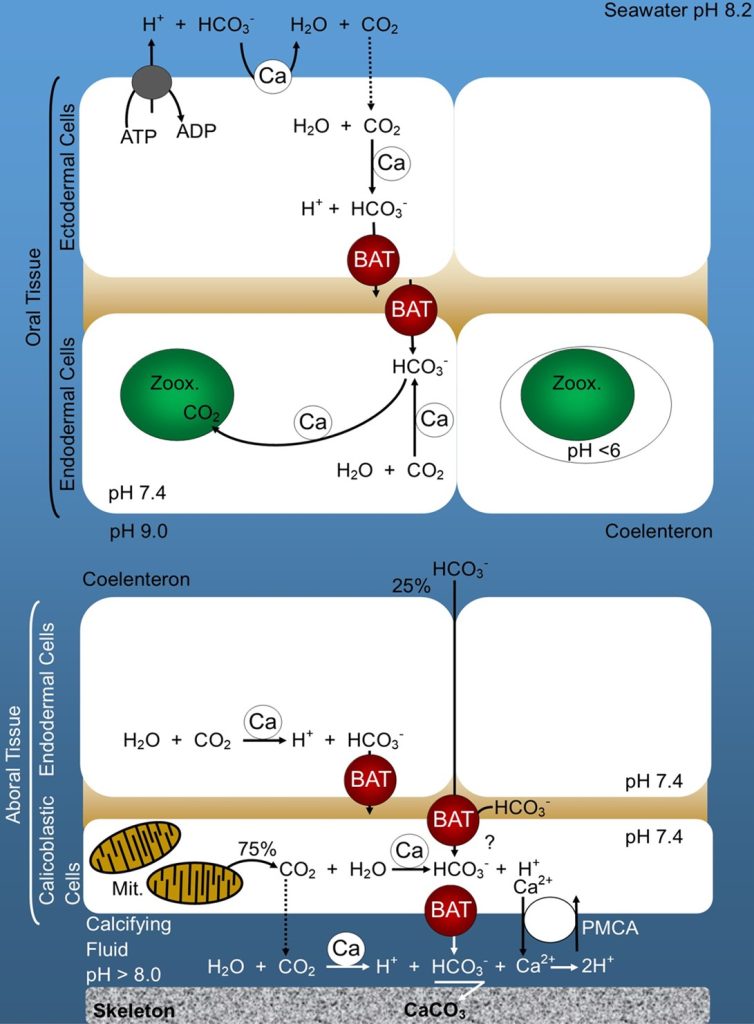
Fig 2. An illustration of coral anatomy featuring the bicarbonate anion transporter (BAT) and the carbonic anhydrase-mediated interconversion of bicarbonate and carbon dioxide (dissolved inorganic carbon; DIC): dotted arrows represent diffusion; [BAT] bicarbonate anion transporter; [Ca] carbonic anhydrase; [Mit.] mitochondria; [PMCA] plasma membrane calcium ATPase; [Zoox.] zooxanthellae; [?] unknown mechanism. Adapted from Zoccola et al. 2015 and Putnam et al. 2017.
Vigorous flow minimizes boundary layers and maximizes photosynthesis which is inversely proportional to fluorescence, so extending lamp emissions into 320 nanometers (nm) UVA can upregulate aesthetic and protective fluorescent proteins at the expense of photosynthesis and perhaps wellbeing (Riddle 2016). Near UVB can be used by zooxanthellae’ chlorophylls a and c2 but it is largely expendable (Zhou et al. 2016; Riddle, personal communication, cited in Neptune Systems 2019) while some far UVB and all UVC remain harmful.

Fig 3. PAR hotspots occurring below some of the earliest LED fixtures at 6” (15 cm) depth expressed in µmol m-2 s-1. These luminaires lacked wide-angle lenses and diffusers, and many colonies bleached or exhibited inexplicable symptoms. Adapted from data presented by Russ in 2013.
A coral’s compensation point is its minimum requirement of photosynthetically active radiation (PAR) whereas photosaturation is a PAR that elicits maximum and unsurpassable photosynthesis beyond which corals must exploit adaptive strategies. Increasing irradiance significantly above saturation leads to photoinhibition which robustly inducts coral and symbiont protective mechanisms including coral innate immunity (Riddle, personal communication, cited in Neptune Systems 2019; Toledo-Hernandez et al. 2023). “Suntan”-analogous melanin and green, cyan, and red fluorescent proteins are critical components of coral defense (Palmer et al. 2010; Toledo-Hernandez et al. 2023).
Compensation points, photo-saturation, and -inhibition vary inter- and intra-specifically and according to symbiont lineage.
Some corals are highly adaptable like those nurturing heat tolerant Durusdinium-like (formerly clade D) Symbiodiniaceae (LaJeunesse et al. 2018) yet many such zooxanthellae support merely decelerated growth (Boilard et al. 2020).
Several mitigating cascades manufacture antioxidants, antimicrobial peptides (AMPs) and fluorescent and reflective proteins (Riddle, personal communication, cited in Neptune Systems 2019). Bacterial symbionts undergo compensatory proliferative and metabolic shifts known as metagenomic plasticity whereas the breakdown of mutualism (dysbiosis; bleaching) is likely instigated by the host which represents an immoderate and necessary immunological response (Palmer 2018; Boilard et al. 2020). Cyanobacteria and some microeukaryotes proliferate to reimburse the coral throughout phases 2. and 3. of microbial purging (Boilard et al. 2020; Bonacolta et al. 2023). Deep endolithic Chlorophytes of the genus Ostreobium (McCauley et al. 2022) lessen harmful skeletal reflectance in bleached colonies (Bonacolta et al. 2023) while corals inhabiting hostile tidal pools may be devoid of zooxanthellae on their upper surfaces (Riddle 2016).
Blue, pink, and purple SPS colonies reflect some useful wavelengths and thus require the most intense light, while many aesthetic compounds are not produced until photosaturation (Riddle, personal communication, cited in Neptune Systems 2019).
Fluorescent tubes and metal halide (MH) high intensity discharge (HID) lamps were the only efficacious lighting solutions for SPS corals. Most enthusiasts still use MHs because they mimic the sun by generating a ball of plasma, their somewhat unidirectional emissions interplay with surface movement to create shimmer lines which strobe across corals in nature, while their photons ricochet to create meaningful up- and side-welling (backscatter) for essential underside and lateral illumination of branching and plating colonies.

Fig 4. PAR hotspots observed at 6” (15 cm) depth with contemporary LED pendants expressed in µmol m-2 s-1. Precipitous PAR attenuation no longer occurs but it has not been eradicated, because diffuser and wide-angle lens technologies have somewhat levelled PAR gradients (Fig 6.). Adapted from data presented by Russ in 2013.
The principal drawback with MH technology is its cost because these fixtures have luminous efficacies of radiation (LERs) of ~76 lumen per watt (lm W-1) making them ~11 percent efficient (Any lamp.co.uk; Lighting.Phillips.co.uk). Several SPS devotees use 400W lamps and fixtures representing a significant dent in operating coffers, because a 400W ballast must draw more to drive the lamp, which require replacing every seven months to safeguard their emissions and most cost between 40 and 50GBP ($51-$64 USD).
Developments in light emitting diode (LED) technology from the tiny red or green dots used as power indicators on our HiFi equipment or cathode ray tube televisions, was meant to provide a real alternative for reef hobbyists. Many of the early LED fixtures generated fatal abnormalities and artefacts that were hard to ignore.
They flooded the light field with damaging radiation immediately beneath at 6 inches depth yet PARs of less than an eighth were observed on the periphery and to the side. Such hotspots presented challenges because corals could not be acclimated for fear of depriving the rest of the display (Fig 3.; Fig 4.)
Furthermore, their emissions left the luminaire as discrete bands corresponding to each LED which created glitterball-like patterns on the surrounding walls which soon became known as “glitterballing”. Most of the radiation could be blended if the lamps were mounted higher, but this severely curtailed their intensity in accordance with the inverse square law.

Fig 5. A healthy “frag” of Acanthastrea species of LPS coral.
“Soft” and large polyp stony (LPS) corals fared better with early generation LED technology albeit Acanthastrea species turned orange and peeled away from their skeleton while tissue loss was observed in Catalaphyllia species (Fig 5.; Russ 2013).
Some anomalies were likely due to the catalytic cleavage of green-to-red fluorescent proteins following exposure to 400 nm radiation (Oswald et al. 2007) insofar as intense blue LEDs are potentially injurious (Riddle, personal communication, cited in Neptune Systems 2019).
Lens and diffuser developments commenced to unify spectra from the fourth and fifth generation. However, LEDs remain unidirectional and somewhat experimental because they lack backscatter, unlike MH and fluorescent lamps. Hence, many hobbyists favor hybrid fixtures incorporating T5 very high output (VHO) fluorescent tubes.

Fig 6. The LED cluster and wide-angle lenses of an EcoTech Radion XR15 4th generation reef fixture.
The author learnt to fabricate his own LED luminaires, the plans of which are illustrated in The Complete Reef Aquarist – A Conservation Manual (Aslett 2024a).
Many hobbyists have reported nil challenges with fourth generation EcoTech Radions that reliably elicit the growth of SPS corals with sufficient underside illumination, which integrate outstanding chips from Cree, Osram, and SemiLED. Nevertheless, the author remains a dedicated fan of HID technology.
Early MH lamp and electromagnetic choke mismatches motivated The American National Standards Institute (ANSI) to classify and inscribe compatible lamps and chokes.
A rare yet meaningful phenomenon called acoustic resonance caused spectral emission disparities, misshaped arcs, and occasional arc tube rupture when electronic ballasts were first used to operate MHs.

Fig 7. A cluster of Giesemann® LED chips embedded in wide-angle lenses nestled within parabolic reflectors exemplifying the importance of spectral unification. Image courtesy of Giesemann ©.
Every object has an eigenfrequency at which it naturally vibrates, and acoustic resonance occurred when lamp eigen- and ballast-frequency matched (Wiberg et al. 1999; Park et al. 2008). Electronic ballasts have comprised microprocessor frequency searching that detects and compensates for acoustic resonance since 2008 (Moo et al. 2008; Park et al. 2008) while they adopt a switching frequency that is 1,000 times that of the supply and are now eminently dimmable.
-
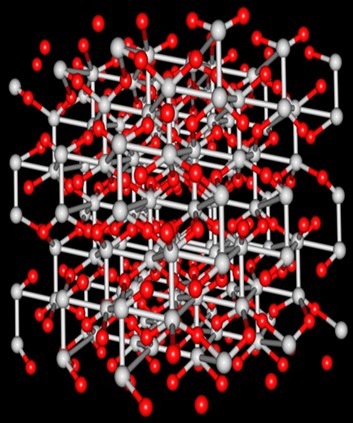
- Fig 8. The crystalline structure of corundum (sapphire; α-alumina) where each aluminium atom (blue) is octahedrally coordinated with six oxygens (red; Liao 2017).
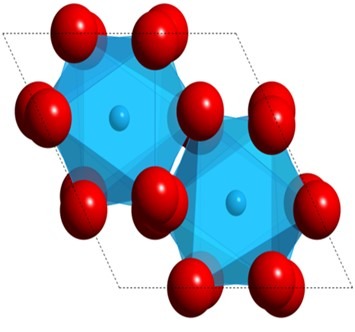
PN-junction LEDs are made from two or three crystalline epitaxial layers, where the second and third veneer adopts the underlying crystal lattice despite their typical configuration. Epitaxial synthesis was devised to minimize imperfections which cannot surpass the substrate’s flawlessness.
LEDs were initially fabricated from sapphire (α-alumina; corundum) which is artificially manufactured by a process called flame fusion from which several types of chip are still made today, yet they have one million imperfections per square millimeter (Fig 8.; Stone-Sundberg 2013).
Diodes prevent the movement of current (electrons) in all but the intended direction so a circuit of several chips wired in series will not illuminate if one is misorientated. The p-type layer has an abundance of inner shells which lack electrons yet its outer orbitals are full, hence these quantum wells can only be filled by electrons with minimal energy. The n-type has a wealth of electrons which travel towards the p-layer where they liberate photons before capture by the degenerate inner shells (Fig 9.; Stone-Sundberg 2013).
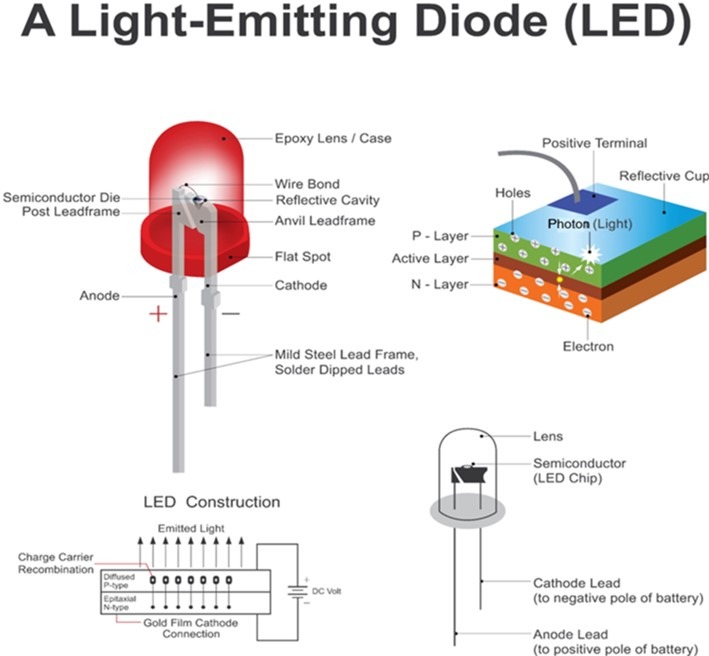
Fig 9. Light emitting diodes (LEDs) are PN-junction diodes which only permit the flow of electrons (current) from n to p.
A glitch in LED development called efficiency droop occurred when early chips were driven at higher currents because heat was generated in their active layers. An inner electron is lost, and photon emission facilitates the accommodation of an outer electron, or energy ejects another electron while much is converted to heat. LED development was thus forestalled before the elucidation of auger excitation and emission (Iveland et al. 2013; Meyaard et al. 2014).
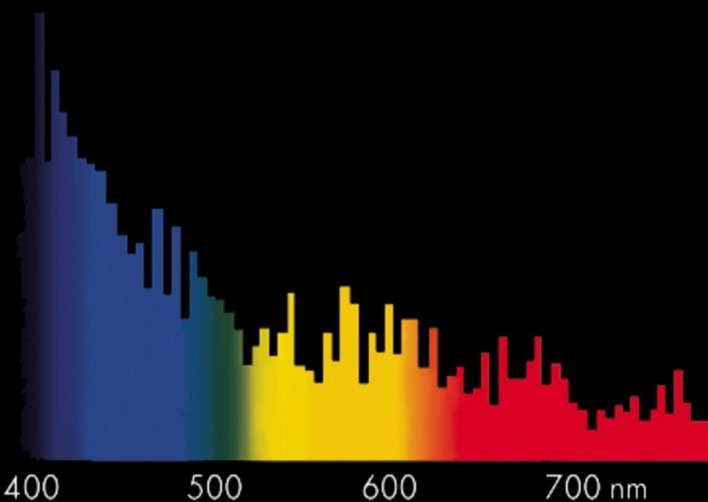
Fig 10. The spectral emissions of a Megachrome Marine double-ended (DE) MH lamp manufactured by Giesemann®. Image courtesy of Giesemann ©.
The metal gallium combines with nitrogen to form the quantum well-abundant and exceedingly flawless semiconductor, gallium nitride (GaN) whereas indium nitride (InN) reacts with GaN to form indium gallium nitride (InGaN). GaN is frequently propagated on sapphire or silicon carbide and overlayed with InGaN.
GaN-on-GaN chips represent a profound yet intuitive leap in LED technology because fabrication is simplified, GaN has one thousand times less imperfections compared with foreign substrates, 15 to 25 percent smaller LEDs emit the same photons, and GaN’s refractive index ensures efficient radiation with nil IR accumulation where trilateral (triangular) chips have improved efficacy (Krames 2013).
Metal halides are formed when a halogen and metal bond covalently (KI) or ionically (Na+Cl–; Busch & Meyers 2003). The ignition sequence of a MH lamp commences when several 5 to 30-kilovolt pulses are delivered to the electrodes where argon is exploited as a starter gas. Streams of energetic electrons bump into gas particles which disrupts some orbitals and induces cationization. Basic atomic structure and ionization are explored in Aslett 2024 (https://www.reefranch.co.uk/). Electrons can only recombine with atoms after they have lost energy in the form of a photon, and thus conductive arc plasma is propagated in a dynamic process whereby electron collisions cause ionization and recapture liberates light.
Rising temperature vaporizes gases during the initial burn which builds slowly and will not startle fish, where arc tube plasma reaches approximately 10 bar (145 pound-force per square inch; lbf in-2; psi; Chammam et al. 2016) and 800 to 1,000oC (1,472 to 1,832oF).

Fig 11. A single-ended (SE) MH lamp with an E39, E40 or the slightly longer EX39 threaded mogul base featuring its arc tube, electrodes, and UV shroud. The envelope of double-ended (DE) lamps is not UVC and B attenuating so they must be concealed behind ordinary glass (Fig 12.).

Fig 12. A 3D parabolic reflector within a MH pendant fitted with a DE lamp with a UV-filtering glass shroud, which must remain intact for fear of irradiating the aquarium with livestock-decimating and sterilizing UVC.
Current and voltage-regulating mercury vapor provides stability for the emission-modulating metal halides and rare earth metals. Yellow, green, blue, and red are created by sodium (Na), thallium (TI), indium (In), and lithium (Li), whereas wavelengths are further influenced by rare dysprosium (Dy), holmium (Ho), and thulium (Tm; Davidson 2015; Osram 2018).

Fig 13. The CoralVue Lumen Bright reflector and pendant for a SE MH lamp was 16.5 percent more efficient than its competitors in 2008. Image courtesy of CoralVue inc. ©.

Fig 14. An SPS coral-dominated reef illuminated with 400W MH lamps focused with computer-designed reflectors. Auxiliary glass shrouds lacking, the results are exemplary. SE mogul lamps are screened with a UV-protective outer envelope.
The author’s extensive experiments indicate that the resolute SPS coral enthusiast will be disappointed with any lighting solution other than metal halide high intensity discharge lamps and their fixtures.
That concludes this instalment; next we will commence to ascertain the basic setup that expedites and sustains the ultra-pure (oligotrophic) borderline nutrient-limited water quality common to pristine wild reefs.
References
Any Lamp BV. Metal Halide Specifications – Osram Powerstar HQI-T 250W D Pro E40, 5,500K Any Lamp BV, Victory House, 400 Pavilion Drive, Northampton Business Park, NN4 7PA Northampton, Northamptonshire. https://www.anylamp.co.uk/
Aslett, C., G. (2024) Reef Digress: Fundamental and Foundational Science: Not Simply Water. Available from: https://www.reefranch.co.uk/
Aslett, C., G. (2024a) The Complete Reef Aquarist, for the hobby, trade and academics – A Conservation Manual. Aslett, C., G. (ed.). Reef Ranch Publishing Ltd. In press. https://www.reefranch.co.uk/
Boilard, A., Dubé, C., E., Gruet, C., Mercière, A., Hernandez-Agreda, A. & Derome, N. (2020) Defining Coral Bleaching as a Microbial Dysbiosis within the Coral Holobiont. Microorganisms. 8, 1682. https://www.doi.org.10.3390/microorganisms8111682
Bonacolta, A., M., Weiler, B., A., Porta‐Fitó, T., Sweet, M., Keeling, P. & del Campo, J. (2023) Beyond the Symbiodiniaceae: diversity and role of microeukaryotic coral symbionts. Coral Reefs. 42, 567–577. https://doi.org/10.1007/s00338-023-02352-0
Busch, M., A. & Meyers, R., A. (2003) Encyclopedia of Physical Science and Technology (Third Edition). Myers, R., A. (ed.). Academic Press, New York. pp 197-200.
Chammam, A., Nsibi, W., Nehdi, M., Mrabet, B. & Sellami, A. (2016) Effects of frequency dimming electronic ballast on a high‐pressure mercury thallium iodide discharge. Journal of the Society for Information Display. 24(3), 154-158.
Davidson, M., W. (2015) Fundamentals of Metal Halide Arc Lamps. Education in Microscopy and Digital Imaging. Zeiss. http://zeisscampus.magnet.fsu.edu/articles/lightsources/metalhalide.html
Houlbrèque, F. & Ferrier‐Pagès, C. (2009) Heterotrophy in Tropical Scleractinian Corals. Biological Reviews. 84(1), 1-17.
Iveland, J., Martinelli, L., Peretti, J., Speck, J. & Weisbuch, C. (2013) Direct Measurement of Auger Electrons Emitted from a Semiconductor Light-Emitting Diode under Electrical Injection: Identification of the Dominant Mechanism for Efficiency Droop. Physical Review Letters. 110(17), 1774061-1774065.
Krames, M. (2013) Light-Emitting Diodes: GaN-on-GaN platform removes cost/performance tradeoffs in LED lighting. Laser Focus World. Soraa, 6500 Kaiser Dr., Fremont, CA 94555. https://www.laserfocusworld.com/articles/print/volume-49/issue-09/features/light-emitting-diodes-gan-on-gan-platform-removes-cost-performance-tradeoffs-in-led-lighting.html
Liao, Y. (2017) Practical Electron Microscopy and Database. Alpha (α)-Alumina (Sapphire). GlobalSino.com. https://www.globalsino.com/EM/page2591.html
McCauley, M., Goulet, T., L., Jackson, C., R. & Loesgen, S. (2022) Systematic review of cnidarian microbiomes reveals insights into the structure, specificity, and fidelity of marine associations. Nature Communications. 14, 4899. https://doi.org/10.1038/s41467-023-39876-6
Meyaard, D., S., Lin, G., B., Cho, J. & Schubert, E., F. (2014) Efficiency droop in gallium indium nitride (GaInN)/gallium nitride (GaN) LEDs. Nitride Semiconductor Light-Emitting Diodes (LEDs), Materials, Technologies and Applications. Huang, J., J., Kuo, H., C. & Shen, S., C. (eds.). Woodhead Publishing. pp 279-300.
Moo, C., S., Huang, C., K. & Yang, C., Y. (2008) Acoustic-Resonance-Free High-Frequency Electronic Ballast for Metal Halide Lamps. IEEE Transactions on Industrial Electronics. 55(10), 3653-3660.
Neptune Systems (2019) #LetsTalkReef – Today its all about measuring light – with Guest Dana Riddle. https://www.youtube.com/watch?v=LKlFA59-Acc
Osram (2018) Metal Halide Lamps, Instruction for the use and Application. https://docplayer.net/20725352-Www-osram-com-metal-halide-lamps-instructions-for-the-use-and-application.html
Oswald, F., Schmitt, F., Leutenegger, A., Ivanchenko, S., D’Angelo, C., Salih, A., Maslakova, S., Bulina, M., Schirmbeck, R., Nienhaus, G., U., Matz, M., V. & Wiedenmann, J. (2007) Contributions of host and symbiont pigments to the coloration of reef corals. The FEBS Journal. 274, 1102-1122.
Palmer, C., V. (2018) Immunity and the coral crisis. Communications Biology. https://dio.org/10.1038/s42003-018-0097-4
Palmer, C., V., Bythell, J., C. & Willis, B., L. (2010) Levels of immunity parameters underpin bleaching and disease susceptibility of reef corals. The FASEB Journal. 24,.
Park, C., Y., Kim, K., N. & Lee, B., J. (2008) High-Power Electronic Ballast Design for Metal-Halide Lamp without Acoustic Resonance. The Transactions of The Korean Institute of Electrical Engineers. 57,.
Philips Lighting Holding B.V. (2000) T12 Specifications – High Tech Campus 45, 5656 AE Eindhoven. http://www.lighting.philips.com/
Putnam, H., Barott, K., Ainsworth, T. & Gates, R. (2017) The Vulnerability and Resilience of Reef-Building Corals. Current Biology. 27(11), 1551-1706.
Riddle, D. (2016) Feature Article: Montipora digitata: A Stony Coral for All Hobbyists. The Marine Aquarium Council of North America. BulkReefSupply.com. https://youtu.be/fBktpJ3umAs [last accessed 16 August 2018].
Russ, K. (2013) Part III of the Paletta LED Experimental Saltwater Reef Tank – with Mike Paletta and Sanjay Joshi. https://www.advancedaquarist.com/blog/part-iii-of-the-paletta-led-experimental-saltwater-reef-tank-with-mike-paletta-and-sanjay-joshi
Stone-Sundberg, J. (2013) Sapphire Series Part 1 to Part III: Introduction to Sapphire and Synthetic Sapphire. The Gemstone Institute of America Inc. https://www.gia.edu/gia-news-research-Sapphire-Series-Introduction-to-Sapphire-and-Synthetic-Sapphire
Toledo-Hernandez, C., Ruiz-Diaz, C., P., Ramirez-Lugo, J., S., Torres-Diaz, M., Santiago-Pagan, L., Bruno-Chardon, A. & Diaz-Vazquez, L., M. (2023) Uncovering the link between environmental factors and coral immunity: A study of fluorescent protein expression and phenoloxidase activity in Acropora cervicornis. Forntiers in Marine Scinence. https://dio.org/10.3389/fmars.2023.1133486
Wiberg, N., Bausys, R. & Hager, P. (1999) Adaptive h -version eigenfrequency analysis. Computers & Structures. 71(5), 565-584.
Zhou, J., Fan, T., Beardall, J. & Gao, K. (2016) Incident Ultraviolet Irradiances Influence Physiology, Development and Settlement of Larva in the Coral Pocillopora damicornis. Photochemistry & Photobiology. 92(2), 293-300.
Zoccola, D., Ganot, P., Bertucci, A., Geminti-Segonds, N., Techer, N., Voolstra, C., R., Aranda, M., Tambutte, E., Allemand, D., Casey, J., R. & Tambutte, S. (2015) Bicarbonate transporters in corals point towards a key step in the evolution of cnidarian calcification. Scientific Reports. 5, 9983.


0 Comments