

Chaetodontoplus caeruleopunctatus from Cebu, Philippines. Credit: brucelee / Aquarius Divers
At first glance, the Bluespotted Angelfish (Chaetodontoplus caeruleopunctatus) is a wholly unremarkable fish, sorely lacking in the intricate patterning and kaleidoscopic colors that one might expect from a pomacanthid. And, unsurprisingly, it tends to garner little interest amongst aquarists, but, what it lack in looks, it more than makes up for in mystery. The species was first discovered by aquarium collectors in the 1970’s from the island of Cebu in the Philippines and, since then, has only been reported from only three more nearby locations: Anilao, Samar, and Zamboanga. But, despite its apparent rarity, this fish has been a consistent offering in the aquarium trade, shipped regularly in small numbers. So how is it that this nearly foot-long angelfish has continued to remain so elusive, with just a handful of photographs documenting its existence in the wild?
This can partly be explained by its choice of habitat, as this species, like most others in its genus, tends to favor moderately deep waters and rocky environments dominated by algae and sponges… that is to say, the kind of ecological niche that recreational divers seldom visit. Several of its congeners have proven to be equally elusive, such as C. vanderloosi, known only from the eastern tip of New Guinea. The breathtaking beautiful Ballina Angelfish—Australia’s answer to Genicanthus personatus—is equally rare, having been found at just two subtropical locations in the Coral Sea. And then there is C. niger, one of the more enigmatic species in the angelfish family, known from just a handful of specimens and still yet to be photographed in its adult coloration.
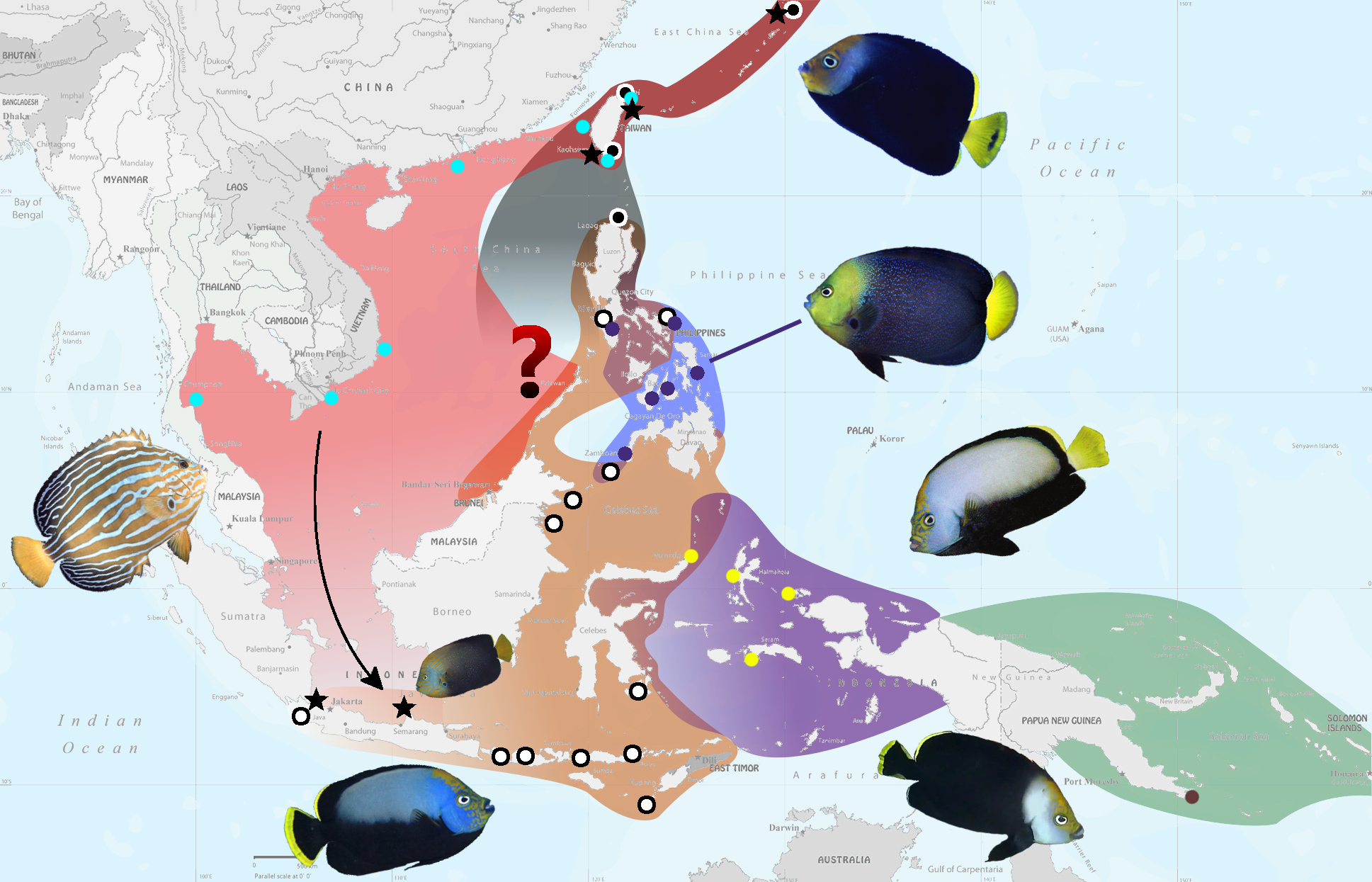
But the greater mystery surrounding C. caeruleopunctatus relates to its unusual distribution, something that only becomes apparent when examining the biogeography of this genus as a whole. It’s endemism in the Philippines would naturally lead us to look for its closest relative further south in Indonesia. This is part of a larger trend observed in Pacific reef fishes—consider the Tomato Anemonefish and Diadem Dottyback (both found in the Philippines), replaced by the Cinnamon Anemonefish and Bicolor Dottyback in Indonesia. Many other examples exist which illustrate the utility of this trend, but, for our angelfish, there is no obvious relative to be found here.
The next logical place to find its sister species would be to the north around the waters of Japan and Taiwan, but, unfortunately, the fishes found here have been inadequately studied, leaving us with an enormously confusing morass of taxonomic uncertainty and phenotypic plasticity. The most distinctive of these is the Bluelined Angelfish (C. septentrionalis), unmistakable thanks to its blue linear patterning. Most sources list this fish as being found only in the Pacific Northwest (Japan, Taiwan, China, Korea), but aquarium specimens have been regularly exported from Vietnam and there are reports from Ko Tao in the Gulf of Thailand, indicating that this species is likely common throughout continental Southeast Asia.
At the other end of the spectrum is a predominantly black fish that I have dubbed elsewhere as the Japanese Velvet Angelfish. This is presumed to be an undescribed species closely related to C. melanosoma, a widespread fish in the Northern Philippines, Northeastern Borneo and south into the Lesser Sunda Islands and Southern Sulawesi. The two have long been conflated due to their nearly identical appearance, but there are some important and consistent differences seen in their coloration, with the latter species having a fully grey back and mostly black caudal fin. Aquarium specimens of this undescribed fish have been commonly collected from the Babuyan Islands at the northern edge of the Philippines, leading some to misidentify them as C. vanderloosi, the rare New Guinean endemic.

Mixed pair, C. septentionalis and a hybrid “chrysocephalus” phenotype. From Izu, Japan. Credit: T. Ueno
Things get even murkier when we introduce the Maze Angelfish (C. cephalareticulatus) and Orangeface Angelfish (C. chrysocephalus) into the equation, two of the rarest and most sought after species in the aquarium trade. Both were scientifically described in 1975 from specimens collected off Northeastern Taiwan, but, until recently, few had been seen in the wild and there has been considerable question about how these two truly differed. Thanks to the efforts of Japanese recreational divers, we now have an exponentially greater number of photographs which have revealed with remarkable clarity just how variable these fishes are—while some specimens can be almost entirely covered in irregular blue lines and reticulations, others seem to lack them almost entirely, appearing mostly grey or black.

Mixed pair, C. septentrionalis and C. cf melanosoma. From Kashiwajima, Japan. Credit: kazkian
What’s more, we now have numerous examples of these fishes occurring in mixed-species pairs with the Blueline Angelfish, seeming to confirm a long-standing hypothesis that these dubious species are merely hybrids. This would of course go a long ways towards explaining their general rarity and phenotypic plasticity, and it has been reported that there is unpublished genetic data to support this contention. The likely parent species are C. septentrionalis and the Japanese Velvet Angelfish, and, importantly, these hybrids help to give us some valuable insight into C. caeruleopunctatus and its distinctive blue markings.

Hybrid phenotype from Kashiwajima, Japan showing fine blue spotting along the sides reminiscent of C. caeruleopunctatus. Credit: kiss2sea
In the most reduced state (perhaps representing a backcross with C. cf melanosoma), these hybrids can possess a fine constellation of blue scales against the slaty coloration of the body, appearing nearly identical to the Bluespotted Angelfish, save for the blue markings present on the forebody. Extrapolating out from this, we can hypothesize that the Blueline Angelfish is likely the closest relative of C. caeruleopunctatus, which is further supported when we compare their mitochondrial CO1 and CytB genes.

Small aquarium specimen of C. caeruleopunctatus from Zamboanga, Philippines. Credit: Barnett Shutman / RVS Fishworld
In a previous discussion of this group, I had suggested that the Bluespotted Angelfish was perhaps just another rare hybrid created when the occasional C. septentrionalis wafted over into the Philippines from its nearby population along the Asian mainland. This makes some intrinsic sense, given how rarely encountered C. caeruleopunctatus is in the wild, but there are also several arguments against this theory. For instance, there are no records of the Blueline Angelfish from anywhere in the Philippines, though there are some unconfirmed reports of the Maze Angelfish having been found. Equally problematic is the relative abundance of specimens collected for the aquarium trade and their fairly homogenous appearance—hybrids are, almost by definition, fairly rare and often quite variable.
Barnett Shutman of the Philippines-based aquarium wholesaler RVS Fishworld reports that specimens are acquired from Zamboanga at the southeast corner of Mindanao, and that divers normally encounter them in single-species pairs. Again, this is not what we would expect to find in a hybrid, though a recent dive video taken from an unnamed location in the Philippines documents a mixed-quartet comprised of three C. melanosoma and a single C. caeruleopunctatus. This likely hints at some amount of interbreeding between these two closely related and similarly patterned fishes. To understand the mysterious origins of the Bluespotted Angelfish, we need to consider what the West Pacific looked like in its recent geologic past.
Were we to travel back in time to the last period of glaciation during the Pleistocene (which only ended around 12,000 years ago), we would find a very different landscape in this region. With much of the earth’s water stored in massive ice sheets, sea levels are estimated to have been 100 meters or more lower than their present condition, with the result being that the the major islands of Indonesia (Sumatra, Java, Borneo) were joined together as a single landmass connected to the Asian mainland, referred to as Sundaland.
For widespread species in the region, this would have had a drastic effect on their ability to disperse and interbreed, creating unique allopatric populations wherever these physical barriers limited gene flow. Over enough generations, these shifting geographic boundaries, brought about by vacillating sea levels, worked to create much of the biodiversity seen in the Indo-Pacific today. This is particularly well-studied for the so-called “geminate species” that formed when the Indian and Pacific Oceans became divided by Sundaland, but much less is known about the effect this had within the Coral Triangle.
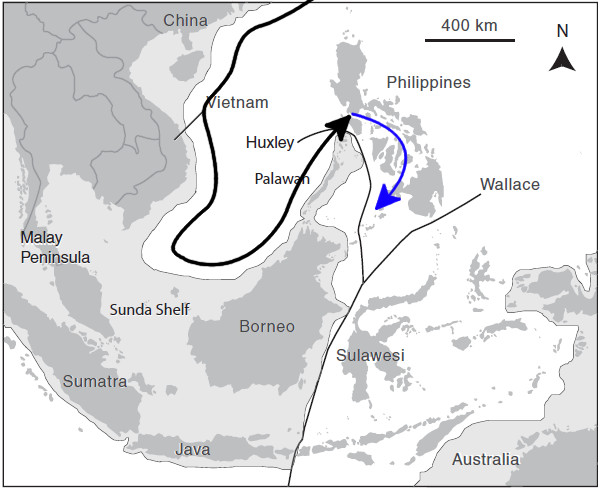
Hypothesized dispersal route of C. septentrionalis (black arrow) into the Central Philippines to hybridize and create C. caeruleopunctatus (blue arrow).
If we look at the geographic boundaries of closely related species within this region, we can get some idea for how speciation progressed. Since Alfred Russel Wallace first pondered this phenomenon in the 1860’s, biogeographers have suggested a number of imaginary lines marking where one fauna stops and another starts. Wallace drew his line through the Celebes Sea and the Makassar Strait, lumping the Philippines along with Sundaland. Another naturalist by the name of Lydekker drew his line around the shared fauna of New Guinea and Australia and, in the process, created a middle region commonly referred to as Wallacea, where Melanesian and Southeast Asian species have been free to intermingle. This rather nebulous area is home to an astounding number of coral reef species (likely a result of this intermixing), including another of the velvet angelfishes, Chaetodontoplus dimidiatus.
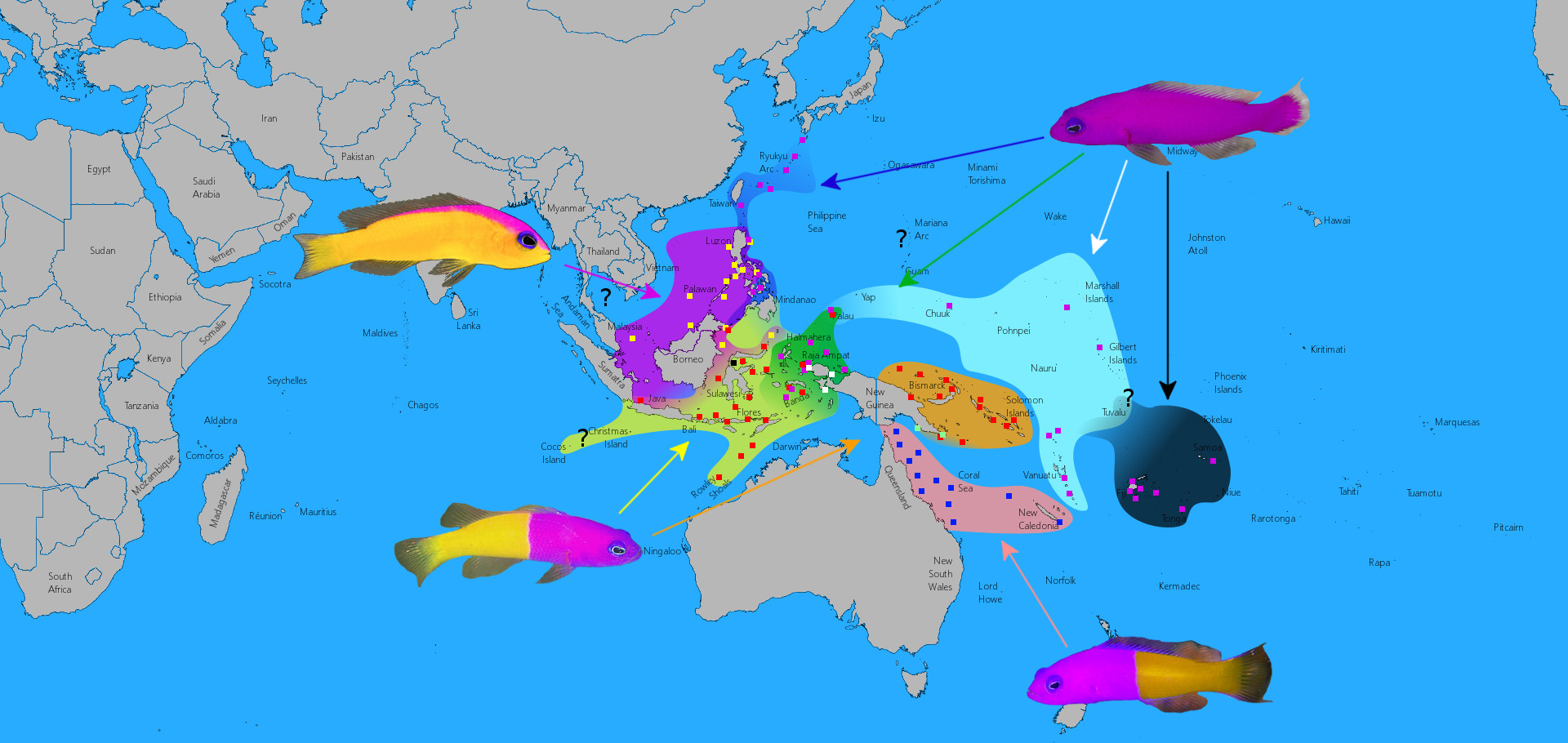
The biogeography of Pictichromis shows a nearly identical pattern of speciation as that seen in the Chaetodontoplus melanosoma group.
When we look at the distribution of C. caeruleopunctatus, we find that it is limited along its southern border by Wallace’s Line. During times of lowered sea levels, a nearly complete land bridge is thought to form from Zamboanga to Borneo, creating an impenetrable barrier that limits dispersal into and out of the Philippines. This has led to speciation in many groups (such as with the Tomato Anemonefishes and the Pictichromis Dottybacks), but many others have crossed this divide without any obvious diversification. This is a fascinating phenomenon that is essentially unstudied. What allows for one group to speciate while another doesn’t? The answer might relate to the relative ages of the clades or some yet unknown factor concerning their ecology or life history, but we essentially know nothing about the details of this mechanism.Knowing that the Blueline and Bluespotted Angelfishes are sister species, we can now envision a route that brought the Philippines its peculiar pomacanthid. During periods when Sundaland was above sea level, there should have been a clear path allowing for colonization, from Malaysia to Borneo and across the Palawan land bridge into Mindoro and the rest of the Philippines. Separating Mindoro and Palawan is a deep turbulent strait which is presumed to form an impassable barrier for both marine and terrestrial organisms, and it is thus only during these periods of glaciation that this region becomes penetrable. This was recognized by yet another biologist, Thomas Henry Huxley, who redrew Wallace’s Line by snaking it across the Sulu Sea and through the Mindoro Strait.
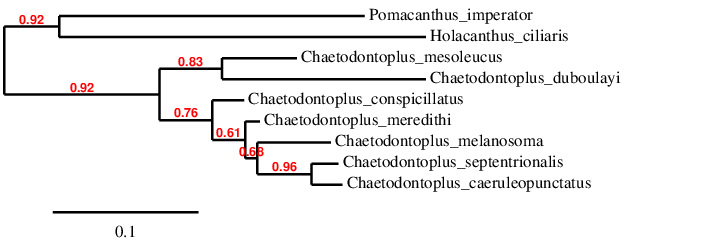
Mitochondrial CytB ML phylogeny, illustrating the close relationship of the C. melanosoma group. Data from GenBank.
Huxley’s Line is critical for understanding the speciation that has occurred in this region among a wide range of organisms. While marine fishes are essentially unstudied in this regard, there is a growing amount of genetic data from other groups illustrating how Palawan has acted as a so-called “biogeographic umbilicus” linking the fauna of Sundaland with the Philippines. One particularly germane example can be observed with the Giant Freshwater Prawn (Macrobrachium rosenbergii), which occurs throughout Sundaland (west to Pakistan) and the Philippines, with a distinct genetic divide showing that the two have been diverging since the Miocene, more than 5 MYA. For the Rana signata complex of stream frogs, both the Palawan and Sulu land bridges acted to foster emigration out of Sundaland and into the Philippines.
As a species associated with the rocky, continental coasts of Southeast Asia, it’s not hard to imagine how a Miocene or Pleistocene ancestor of the Blueline Angelfish would have acted just like these freshwater prawns and stream frogs, jumping across land bridges to reach a new home. Once these bridges closed with the rising of sea levels, the population left behind would have been genetically isolated and likely outnumbered by the local fauna. In this instance, the ancestral C. caeruleopunctatus likely interbred with C. melanosoma, creating hybrids similar to what has occurred with the Maze Angelfishes, and eventually leading to speciation. Without a resident population of pure-blooded Bluelined Angelfishes, this miscegenated Filipino fish was free to follow its own evolutionary bent.
This phenomenon is likely to be widespread among coral reef fishes, especially in areas where distinct regional faunae overlap, but only recently have marine biologists begun to investigate this unique form of speciation. Needless to say, their is a great deal of research needed to confirm what I’ve proposed here. A large-scale population-level study is required to sort out the finer details, though this is made all the more difficult given that we still don’t know the full extent of their populations. The northern coasts of Borneo and Palawan, in particular, have seen little exploration and development, with the result being that we know virtually nothing of the fishes found there. While these regions are likely depauperate in lush coral habitats, they presumably abound in the sort of rocky coasts that these fishes call home. Either one of these angelfishes could be expected to occur there, with the possibility that they are still actively interbreeding and hybridizing not outside the realm of possibility.
Though the Bluespotted Angelfish may not be winning any beauty contests, it is undeniably a fascinating and unique coral reef fish. It’s unusual evolutionary origins may one day serve as an important model for the process of hybrid speciation and the important role this likely plays in creating new biodiversity on Indo-Pacific coral reefs.
- Allen, G. R. and M. V. Erdmann. 2012. Reef fishes of the East Indies. Volumes I–III. Tropical Reef Research, Perth Australia. v. II: 425–855.
- Brown, R. M. and S. I. Guttman. 2002. Phylogenetic systematics of the Rana signata complex of Philippine and Bornean stream frogs: reconsideration of Huxley’s modification of Wallace’s Line at the Oriental–Australian faunal zone interface. Biol. J. Linn. Soc. 76(3): 393–461
- de Bruyn, M., JWilson, J. A. and P. B. Mather. 2004. Huxley’s line demarcates extensive genetic divergence between eastern and western forms of the giant freshwater prawn, Macrobrachium rosenbergii. Mol. Phylogenet. Evol. 30: 251–257
- Esselstyn, J. A., Oliveros, C. H., Moyle, R. G., Townsend Peterson, A., McGuire, J. A. and R. M. Brown. 2010. Integrating phylogenetic and taxonomic evidence illuminates complex biogeographic patterns along Huxley’s modification of Wallace’s Line. J. Biogeogr. 37: 2054–2066
- Rowlett, J. 2015. A Look at the Chaetodontoplus. Reefs Magazine. Retrieved from https://reefs.com/magazine/a-look-at-the-chaetodontoplus-178.html/
















0 Comments