

By Lemon TeaYK and Joe Rowlett

The biogeography of the Roaops subgenus.
The butterflyfishes of the family Chaetodontidae are undoubtedly some of the ocean’s most charming and charismatic reef inhabitants. Although entirely unrelated to the terrestrial insects that share the same name, the moniker it bears is highly befitting for these largely shallow-water, coral garden perusing fishes. The family attains maximum diversity in the warm, nutrient-rich waters of the Pacific and Indian Oceans, although they are notably well-documented in the Atlantic as well. Chaetodontidae is highly diverse, with a plethora of species conquering nearly all habitats and niches. From waist deep, sun speckled Acropora gardens, to bone- crushing dropoffs well into the mesophotic twilight zone, there are butterflyfishes to be found in almost every habitat.
Taxonomically, the family is split into ten genera, with the most diverse of these (Chaetodon) being further split into numerous subgenera. Roaops is a small subgenus composed of five handsome and extremely dapper species, which has at times been treated as a full genus (Nalbant, 1984). Genetic data (Fessler et al 2007) has argues against this, however, as these five fishes appear to belong to a larger clade of Chaetodon, including familiar species like C. miliaris and C. citrinellus. Still, Roaops does form a cohesive and recognizable group, identifiable by the following shared traits.
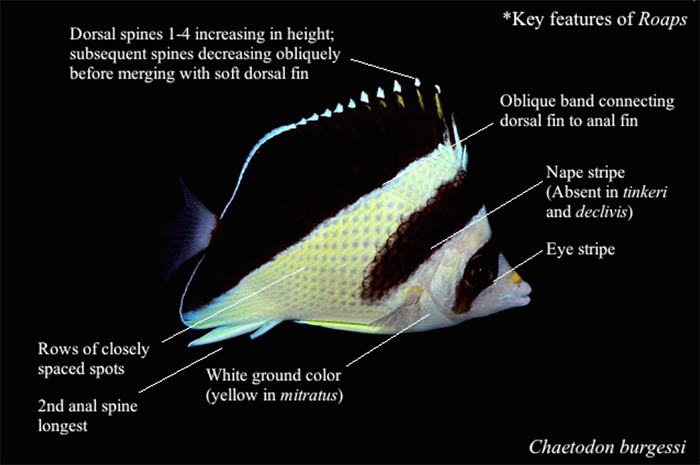
1) The dorsal spines are high and strong, increasing in size from the first to fourth, and then decreasing progressively together with the margin of the soft dorsal. The second spine of the anal fin is also very long and sturdy. These features are also found in Amphichaetodon, Prognathodes and Roa; in fact, the name Roaops translates as “Roa-like” because of the shared morphological traits. However, genetic study indicates that these four groups are only distantly related and their similarities are likely a result of convergent evolution.
2) The scales of Roaops are large and disposed in subhorizontal rows below the lateral line and obliquely upward above the lateral line. This scale pattern can be seen clearly in the manifestation of the body spots. Below the lateral line, these spots are subhorizontal, while above the lateral line, these are oblique. The pattern can be seen more clearly in the individual species profiles later, especially those of C. tinkeri. This is different from Roa and Amphichaetodon, where the scales are small, uniform and arranged in horizontal rows.
3) Meristically, Roaops differ from Roa in having thirteen versus eleven dorsal spines. The number of perforated scales of the lateral line also differ, being more than 42 in Roa and less than 40 in Roaops.
4) In terms of coloration, Roaops are some of the most distinctively patterned in the family. Their design is largely based on vertical or oblique bands; a characteristic shared loosely with certain members in the genus Prognathodes.

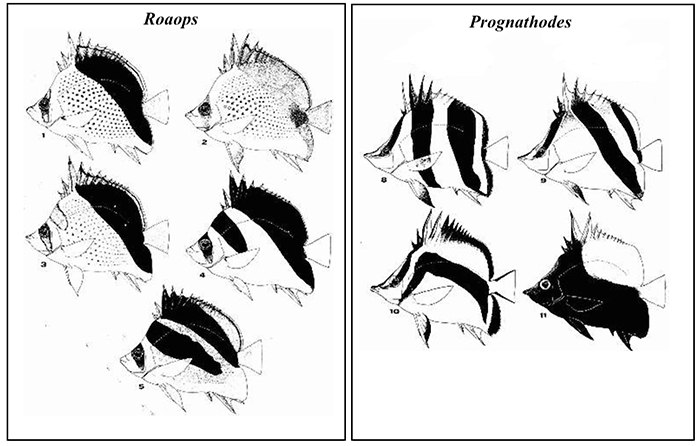
Comparison in oblique banding between Roaops and Prognathodes. Photo by Teodor T. Nalbant.
The biogeography of Roaops is somewhat aberrant for such a widespread group. While these fishes range from Africa to Hawaii, they are curiously absent from certain locations where we would expect to find them, such as Japan and the French Polynesia. The absence from the latter region is particularly striking given the presence of a population at the isolated Marquesas Islands in Polynesia, likely indicating a colonization from the nearby Line Islands to the north.
All are strictly deepwater, but, in spite of this, they are very well known in the aquarium hobby. The Roaops are some of the hardiest and most voracious butterflyfishes, making them suitable for aquarists with virgin track records in this family. Because of their omnivorous benthic diet, they are able to adapt easily to a wide range of prepared foods. The only heartbreak is that these often come with a hefty price tag.
The phylogeny for this group can be broken down into three distinct lineages: Indian Ocean, West Pacific/Micronesia, and Polynesian. It’s tempting to infer relationships between these, but there is no way of knowing what the true evolutionary relationships are until a thorough genetic study is undertaken across the Indo-Pacific. The trees presented below are the two most likely scenarios. Note that “flavocoronatus” is treated as a likely hybrid in this study and may not represent a true “biological species”.
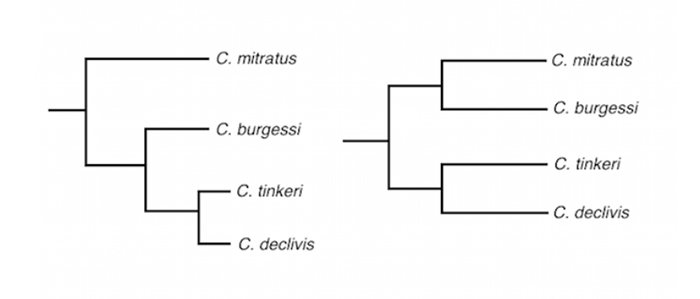
Two proposed phylogenetic relationships of the Roaops subgenus. In the first tree on the left, C. tinkeri and C. declivis are hypothesized as evolutionary derivatives of C. burgessi, which in turn is hypothesized to be the geminate sister of C. mitratus. Although this seems more likely, we cannot be sure if burgessi is more closely related to tinkeri/declivis or mitratus.
The mitratus Clade
Both C. mitratus and C. burgessi are notable for having an exaggerated forehead band. They differ from the tinkeri clade in possessing oblique eye bands, which will serve as an important indication for hybrid diagnostics in the later part of this article. The two are most likely geminate twins, with large geographical distributions within their respective oceans. They probably overlap very sparsely along the western border of the Indonesian archipelago where the two oceans meet, and if so, it is likely that they both hybridize within this narrow contact zone.
Chaetodon mitratus

Chaetodon tinkeri swimming in situ. Photographed in the Maldives. Photo by Maldivian Stars.
Arguably one of the most beautiful and beguiling of the butterflyfishes, C. mitratus, with its highly contrasting blend of rich saffron and black exudes an enigmatic mystery and curiosity, not unlike a shadowy penumbra conjured from the fading sun. In this species, the ground coloration is a saturated, luxurious saffron throughout. The epithet “mitratus” translates to turban, which is rather aptly coined for the copious oblique banding present near its nape. Its eye is concealed in a jet-black band starting at the base of its forehead, extending to the start of the operculum. A second oblique band runs at a 45° angle behind the first, where it stops just at the precipice of the anal fin. A third, much thicker band follows closely behind, engulfing the entire back and dorsal fin, save a narrow margin at the posterior soft rays, where it is orange. A small black ocellus is usually present within this orange boundary, and, at least in juveniles, is very clearly delimited. The ocellus tends to fade with age, but adults are not uncommonly seen with a vestigial remnant in the form of a black smudge. The perimeter of all its bands are lined in a white or light blue, and a series of horizontal stripes comprising finely spaced bluish spots are present across its lower body. The pelvic, anal and caudal fins are yellow, but, in the latter, a thick submarginal hyaline portion is present. All median fins are unmarked.
C. mitratus is vigorously distributed across the Indian Ocean, tracing the East African coast in its westernmost limit. It ranges eastwards to Seychelles, Mauritius, Maldives and the Chagos Archipelago. C. mitratus is sparsely known from the Indian Ocean side of the Indonesian and Thailand borders, and, although present, its prevalence here is poorly documented. It ranges as far east to the Andaman Sea and Jakarta, and attaints its easternmost limit at the Cocos-Keeling and Christmas Island; although there is little doubt that this might extend further to Bali. Should it range to Bali, it puts it in close contact with its sister species, Chaetodon burgessi. However, whether or not the two form hybrids is unknown, as none have been documented yet.

Chaetodon mitratus photographed in Koh Bon, Thailand. Photo by Kairyu Diving.
This species, as with all Roaops, is a deepwater specialist and rarely found at depths shallower than 45m. It comfortably swims well into the mesophotic zones at depths hitting 60-80m. Unsurprisingly, its habitat consists of detritus laden dropoffs and steep walls, where it swims playfully upside-down or sideways, with its lower body hugging the reef catacombs closely. Non-photosynthetic corals such as Tubastraea and various gorgonians take precedence here. Despite its depressing preference for gloomy deepwater reefs, this species remains unfazed by the many hundred watts of light blasted onto it by coral aquarists.
C. mitratus gives very little care to its captive environment, and will thrive in most decent set ups. Apart from the typical butterflyfish penchant for Cryptocaryon, this species feeds well and adapts readily to the captive environment. The drawback? It is uncommon and expensive. But for such a magical pet, that is a small price to pay.
Chaetodon burgessi

A creamy morph of C. burgessi swimming in its habitat at the Togean Islands. Photo by Massimo Boyer.
This species is most similar to C. mitratus in possessing the same exaggerated oblique bands, which gives us ample reason to suspect that the two share a common ancestor which presumably speciated during periods of lowered sea levels that acted to isolate the oceanic basins. What is needed, however, is more genetic sequencing to allow for correlating dates to the genetic divergence, which would give us a better understanding for which glaciation event began their split. In all likelihood, this was not caused by a single event, but a series of them spread over the last 2-3 million years.
In the variable C. burgessi, the ground coloration, like desserts from a French pâtisserie, ranges from Chantilly white to a creamy zabaglione. The eye is concealed in an oblique black band which connects at the top to form a bonnet. Another oblique band follows closely behind, making a more acute angle downwards as compared to the gentler 45° slope of C. mitratus. This band ends behind the pectoral fin base. The back is featureless and black, much like C. tinkeri. The soft dorsal fin is edged thinly in white. Both pelvic fins and pectoral fin bases are tinted in yellow, and a small yellow spot is often present at the base of its rostrum. The body is decorated in the same pattern of linearly placed spots, but this is inconsistent and can vary from strongly to barely present in both color forms. It is also interesting to note that the yellowish, creamy form of C. burgessi does not range into Micronesia and apparently reaches its limit at West Papua.

Highly xanthic specimens of C. burgessi from the Philippines. Note the variability in the body spotting of the two specimens. Photo by OceanandSea.

A pair of C. burgessi from the Coral Sea. This region presents a new range extension for this widespread species. Photo by Rob Lanceley, Cairns Marine.
The white form is found throughout its range and overlaps with the creamy forms in the Western Pacific. It is possible, although at this point completely hypothetical, that the yellowness could be an influence from C. mitratus. Immediately past West Papua in the Coral Sea region to Micronesia, the regional, biogeographic hotspots for endemism changes, and the flow of C. mitratus gene is disconnected, resulting in the absence of yellow specimens. This is of course quite contentious and hypothetical, and absolutely no confirmation on the subject can be reached without genetic testing.

C. burgessi is the most widely distributed of the Pacific Ocean Roaops, and is found throughout Indonesia, Philippines and north to Okinawa. It spreads eastwards to Palau, Guam, Papua, the entire Melanesia and parts of Micronesia. This species was recently documented from the Coral Sea. It straddles the Indian Ocean divide, and is found in Bali, south to Rowley Shoals in Northwestern Australia.
Interestingly, Chaetodon burgessi is exceedingly rare north of the Philippines, and records from Okinawa are the only indication of this species reaching the periphery of Japan. It appears to be absent from the mainland. Like in Japan, its rarity in Guam is also curious, but its confirmed presence here is important with regards to C. flavocoronatus. This will be discussed in greater detail later on.

Chaetodon burgessi swimming in its deepwater habitat in Palau accompanied by Pseudanthias calloura and Pseudanthias smithvanizi. Photo by Umioniyome.
C. burgessi hybridizes intensively with all the other sympatric Roaops to form a plethora of hybrid intermediates. This species is fond of deepwater drop offs and steep slopes in excess of 60m. Unusually, and no one really knows why, it is documented to occur in depths as shallow as 20m in Cenderawasih Bay, where it is often accompanied by other deepwater species, such as Genicanthus bellus.
The species is moderately common in the trade, but it is never abundant. It is the most affordable of the Roaops species and is likewise very hardy and resilient.

Chaetodon burgessi in situ. Photo by Massimo Boyer.

C. burgessi from the Coral Sea. This region represents a new range extension for this wide spread species. Photo by Rob Lanceley, Cairns Marine.
The tinkeri clade
Moving into the Pacific, we find ourselves with three “species” sharing the same overall template of white and black (although in C. declivis, this is somewhat untrue). The similarities between each species is without a doubt a throwback to their evolutionary lineage, and, unsurprisingly, a hodgepodge of intermediates and presumed hybrids run amok in several overlapping regions. The rarely seen and poorly known Chaetodon flavocoronatus is the defining example of contentious species allocation, and we will discuss why this is so later on in this article.
The tinkeri clade can be differentiated from the previous lineage as follows: 1) The caudal fins are translucent yellow in all species. 2) The eye band is always vertically straight, versus oblique in the mitratus clade. This feature makes an exception in C. flavocoronatus, serving as supporting evidence that C. flavocoronatus might be of hybrid origin. 3) Black-spotted scales are present throughout the body, which are separated into two regions above and below the lateral line. This is only weakly and variably present in the mitratus clade.
Chaetodon tinkeri

Chaetodon tinkeri. A stunning example of the Roaops subgenus. Notice the vertical eye band. Photo by Lemon TYK.
Chaetodon tinkeri is another species of exquisite perfection. In this simple but fetching butterflyfish, the ground coloration is alabaster. An oblique sash starting at the second or third dorsal spine to the anal fin edge delimits the back in a featureless black. The edge of its dorsal fin is trimmed in yellow, although the extent is subject to variability, and an eye stripe of similar shade is present vertically from its nape to its operculum. This stripe is present on both sides and they rarely, if ever, conjoin at the upper or lower limits. Much of the anal fin and both pelvic fins are white, while the caudal fin is translucent yellow throughout.
The body is heavily punctuated with fine spots broken up into two regions corresponding to the scale formation. The region below the lateral line is decorated in a series of black spots arranged in neat subhorizontal stripes, decreasing in size towards the posterior. Above the lateral line, another series of spots is present in an oblique fashion.

Chaetodon tinkeri. The haphazardly arranged spots above the lateral line is clearly seen here. Photo by Lemon TYK.
Chaetodon tinkeri has a restricted range and is found predominantly in the Hawaiian Islands, following the archipelago northwards to at least Midway Island. The only other known population is at Johnston Atoll, located southeast of Hawaii roughly halfway towards the Marshall Islands. Outside of this Hawaiian ecoregion, C. tinkeri is highly irregular, with waifs recorded from the Mariana Arc. There, they form hybrids with C. burgessi.
Similar looking hybrids are also documented in Tarawa, Kiribati and Majuro, Marshall Islands. Although these strongly suggest the genetic influence of C. tinkeri, the species has not been reliably documented there. C. declivis, however, has, and because the species is essentially identical to C. tinkeri, hybridization with C. burgessi will likely produce similar looking hybrids, causing the misconstrued idea that C. tinkeri ranges to Micronesia. Again, we will delve more into this later in greater detail.
Like the other Roaops, C. tinkeri is deepwater, and often found in depths exceeding 60m. They adopt the usual playful behavior of swimming upside-down, often in pairs under ledges and steep reef walls. In the cooler waters of Northwestern Hawaii, this species can be encountered, although exceedingly rarely, in shallow SCUBA diving depths. This is consistent with a few other deepwater species, and it is only here where the fabled Genicanthus personatus and Bodianus sanguineus can be seen frolicking about Acropora and stony coral; a highly atypical combination of heaven and earth.
C. tinkeri is not fussy in captivity, and it takes prepared food with gusto. Although never rare, this species cannot be considered common. Juveniles are less frequently encountered, and as with most deepwater fishes, adopt an inverse price scale (i.e, they are more expensive than the adults). This species is the most expensive of the obtainable Roaops.
Chaetodon declivis

Chaetodon declivis from the Marquesan Islands. Note the heavy spotting here. Photo by Luiz Rocha.
C. declivis is highly similar to its Hawaiian cousin C. tinkeri, but it can be readily identified by its variably colored back, which comes in —pun intended— fifty shades of brown. This is the result of its yellow dorsal base coloration mixing with a darker, fuliginous pattern, forming a dirty, smudged appearance. This species is never entirely yellow, with even near perfect specimens showing at least some hint of this infuscation. The scale spots are likewise variable, showing a clinal decrease in prominence as its population nears the equator. The pelvic fins are white or yellow- tinted, and the posterior anal fin is always stained yellow.
This variability has led taxonomists to believe that two separate subspecies are present, the darker ssp. wilderi vs the lighter ssp. declivis. In a metaphorical analogy, tinkeri would be “black haired”, ssp. declivis the brunette, and ssp. wilderi the ditsy blonde. These subspecies allocations suffer the unfortunate fate of being stuck in taxonomic limbo, with various literatures regarding them synonymous, while others treat them as legitimate subspecies. Recent genetic studies have repeatedly found that Marquesan populations of widespread species usually show an amount of divergence worthy of species recognition. So, despite their considerable phenotypic similarties, the Marquesan populations of declivis have a great likelihood of eventually being recognized as a separate species.

C. declivis with a yellower back. Notice the less prominent spotting here, and the distinct yellowing of the anal fin. This serves as a reliable diagnostic separating it from C. tinkeri. Photo by Kaisuigyo.

Chaetodon declivis showing the variability in its yellow coloration. Photo by Lemon TYK.

C. declivis with a comparatively darker, more suffused black back. Photo by Bbox Aquarium.

Chaetodon declivis with a predominantly dark back. Notice the yellow blotch on the posterior anal fin that is always present in this species. Photo by Ohm Pavaphon.

C. declivis showing variations of both subspecific forms. This individual sports a suffused black back with a yellower anterior portion. Photo by Charm.jp.
C. declivis ranges across the Line and Phoenix Islands, and east to the isolated Marquesas Islands. This species waifs to Tarawa and Kwajalein, where it appears to hybridize with C. burgessi to form a confusing variation of phenotypes, which is discussed in greater detail below. There is little doubt that this species also hybridizes with C. burgessi, or at least in part, by creating backcrosses.
The Line Islands and Marquesas are not places frequented by divers and fish photographers, and, because of C. declivis’ range, few photos of this species exist in situ. Nonetheless, this species can be expected to occur in the same habitats as others in its subgenus. This species is common in the trade, although never abundant, and is roughly half the price of C. tinkeri. Aquarium specimens originate from Kiritimati, where both wilderi and declivis forms with varying backs occur. The much yellower wilderi forms are less common.

A particularly black C. declivis. The yellow tinted pelvic and anal fin distinguishes this from C. tinkeri.

Chaetodon declivis in the Marshall Islands. Photo by Kagii Yasuaki.
Chaetodon flavocoronatus and hybridization of the Roaops

A perfect representation of Chaetodon flavocoronatus from Saipan. Photo by Acetomato.
Chaetodon flavocoronatus is perhaps the most enigmatic and inscrutable Roaops. This immaculate beauty is marked similarly to C. tinkeri, with a few important modifications that feature traits from both Roaops clades. It possesses characteristics from C. tinkeri, such as yellowing of the dorsal fin margins and spotted body pattern. In addition, it sports an oblique band, as with C. burgessi, but here it is yellow instead of black. The epithet “flavocoronatus” is derived from this feature and literally translates to “yellow crown”. The eye band is yellow, as with C. tinkeri, but shows slight modifications in its placement. In C. tinkeri, this eye band is always vertical and is rarely connected at the top and lower ends. In C. flavocoronatus, this eye band is slanted ever so slightly toward the operculum and is usually well connected at the top. A yellow spot is present on the upper lip, as with C. tinkeri.
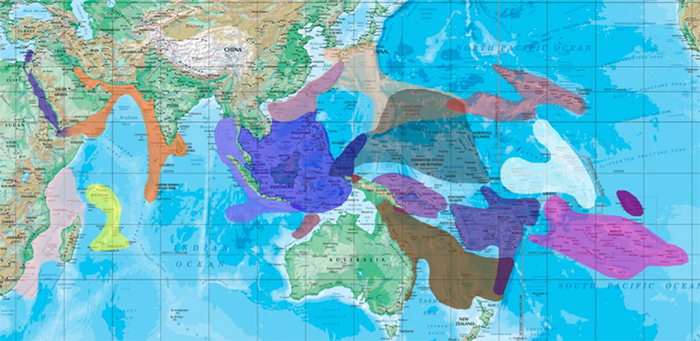
The various subregions of the Indian and Pacific Ocean that harbor high number of endemics, many of which contain sister species of the same group.
In examining the Chaetodon flavocoronatus phenotype, we see an aberrant and bizarre geographical distribution that does not appear to correlate with the usual pattern of allopatric speciation seen in most reef fishes. Instead, this morphologically ambiguous species occupies an equally ambiguous distribution centered primarily on an arc of islands which straddles the ranges of the West Pacific, Micronesian and Hawaiian ecoregions. The evolutionary uniqueness and significance of this region is evidenced in several other groups of reef fishes (e.g. Cirrhilabrus, Genicanthus and Centropyge), all of which appear to have incurred speciation through an intermixing of these distinct geographic populations.

The proposed origin of the flavocoronatus phenotype, as well as the creation of various intermediates. Notice the phenotypic plasticitiy in the Micronesian examples, which will be explained in greater detail to follow.
Chaetodon flavocoronatus is documented at Guam and Saipan in the Mariana Arc, as well as selected regions in the Marshall Islands in Micronesia. This is salient in understanding the hybrid formation of flavocoronatus for two reasons. 1) The Marianas and Micronesia receive different species of Roaops as waifs, which subsequently affects the parental lineages of flavocoronatus in both regions. 2) The ability of the flavocoronatus hybrids to flourish and speciate into a legitimate taxa is very much determined by where it is formed. For example, reproductive isolation in the Mariana Arc is favored over Micronesia due to the low abundance of either of the parental species here.
It’s not entirely possible to solve or speculate the flavoroconatus conundrum without extensive genetic study. What we can do, however, is present the reader with various hypothesis and potential scenarios that may help in deciphering its possible parental lineages.
C. flavocoronatus – Micronesia (Tarawa, Kiribati & Majuro, Marshall Islands)

Chaetodon flavocoronatus in captivity from the Marshall Islands.
It has been long suggested that here, in Tarawa, four members of the Roaops are present. However, only three have been documented with complete certainty — C. burgessi as part of its natural range, C. declivis as waifs from the Line Islands, and C. flavocoronatus as a hybrid derived phenotype. Aside from those, a whole spectrum of hybrids with intermediate phenotypes have also been documented. Because some of these phenotypes bear strong resemblances to C. tinkeri, the presence of the latter in Micronesia has never been doubted.
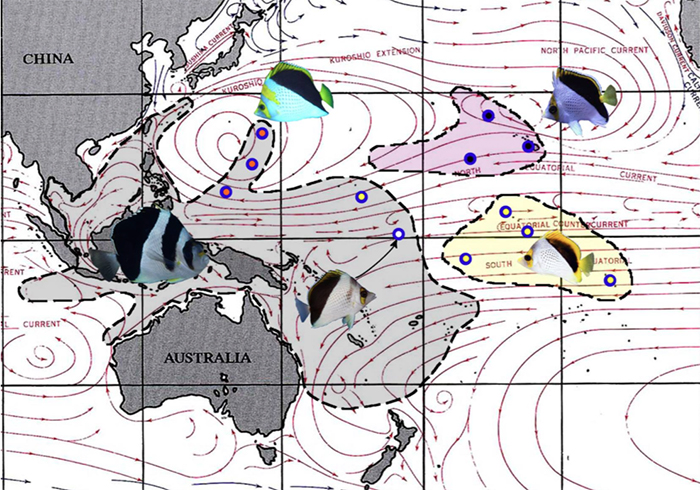
A distribution map of the Roaops butterflies showing the regional surface currents responsible for C. tinkeri and C. declivis being found in Guam and Tarawa, respectively.
The distribution map above showing the regional currents would account for the presence of C. tinkeri and C. declivis in Guam and Tarawa, respectively. Adults aren’t likely to be capable of crossing these vast oceanic distances, but the same is not true of their pelagic larvae. In a study of oceanic currents offshore of Hawaii, (Lobel & Robinson 2002) found that eddies of water supportive of larval fish transport could provide a plausible mechanism for long-distance dispersal. Additionally, major oscillations in the regional currents associated with cyclonic activity and El Nino/La Nina oceanographic patterns may also encourage gene flow.
The direction in current movements would likely explain why after numerous regional surveys conducted by divers and ichthyologists alike, pedigree specimens of C. tinkeri have never been documented in Micronesia. The tinkeri looking hybrids could likely instead come from C. declivis x C. burgessi. Because C. declivis is essentially nearly identical to C. tinkeri, it is entirely possible that the resulting hybrid could be mistaken for those of the latter. Although the exact parentage cannot be determined, it is quite obvious that this orgy of C. burgessi and C. declivis crossing and backcrossing amongst themselves is responsible for the so-called Tarawan “flavocoronatus.” With so many sympatric hybrids and species here in the Marshall Islands, it is unlikely that the flavocoronatus phenotype can attain enough reproductive isolation needed to form a distinct biological species.
C. flavocoronatus – Mariana Arc (Guam + Saipan)

Chaetodon flavocoronatus photographed in situ, Saipan.
The Mariana Arc is known to receive fishes from Japan and Micronesia, as well as a small number of Hawaiian taxa. The distribution of Roaops species here is peculiar, with only three phenotypes found, of which, flavocoronatus is the most abundant. C. burgessi occurs here as a rarity, indicating this region is only weakly connected to the rest of its widespread Pacific Population. Likewise, C. tinkeri makes the occasional appearance as larval waifs from Hawaii. Like in the Marshall Islands, the two species hybridize to form a range of intermediates, one of which is the flavocoronatus phenotype. Again, the exact parental lineage cannot be determined, and likewise it is unknown if a direct F1 hybrid of the two pedigree species is enough to produce the flavocoronatus phenotype. What we do know is that C. declivis never reaches the Mariana Arc, while C. tinkeri does. Therefore although we cannot conclusively confirm the parentage, we can be sure that the Mariana Arc and Marshall Islands phenotype are formed from a mixture of different species respectively.
The rarity of both C. burgessi and C. tinkeri in the Mariana Arc creates a conducive environment for the hybrid derived flavocoronatus phenotype to achieve some form of reproductive isolation. Like many examples in the Mariana Arc, it is likely that this phenotype, given time, will eventually lead to complete speciation, if it hasn’t done so already. This would also explain the relative abundance of C. flavocoronatus here compared to the Marshall Islands, as well as the general lack of phenotypic plasticity.
The whole situation could be thought of as a metaphorical cake recipe. In both locations, similar ingredients are provided to make the perfect cake. The ratio of ingredients in Guam provides a better cake recipe with a greater success rate for perfection. In Tarawa, the ingredients are skewed and disproportionate, lowering the probability and success rate for a good bake.

Centropyge shepardi and Centropyge loricula x C. ferrugata. The similarities in the two foreshadows the hybrid origin of C. shepardi. Photos by Lemon TYK and ReefBuilders.
This hypothesized hybrid origin for C. flavocoronatus in the Mariana Arc is paralleled by numerous other unrelated groups, such as Cirrhilabrus katherinae and Centropyge shepardi (Rowlett, in press). In the latter example, a nearly identical fish is known to coccur in the Philippines from the hybridization of the Flame and Rusty Angelfishes. Centropyge ferrugata is unknown from Guam, but it can be seen as analogous to the equally rare tinkeri in its formation of the shepardi hybrid-species.
Dirty Hybrids

A probable hybrid of C. burgessi x C. declivis, or an F1 x C. burgessi or C. declivis backcross in Pohnpei. Photo by Brian Greene.
Apart from the flavocoronatus phenotype, a whole plethora of hybrids occur throughout all the overlapping regions in the Mariana Arc as well as Micronesia. These are often called “dirty hybrids”, in reference to their dusky or smudged region near the nape. This incomplete crown is caused by C. burgessi, which occurs throughout. And because these hybrids are often fertile, they create an inscrutable spectrum of phenotypes ranging in variability. Likewise, determining the parentage of these hybrids is impossible, but, as with the flavocoronatus phenotype, we are able to at least tell which species are involved based on the biogeography.

Records of C. tinkeri in the Marshall Islands may have stemmed from hybrids such as these. However, upon closer scrutiny, notice that the eye band is oblique and the nape is suffused with black. Photo by In-Depth Images Kwajalein.

Another Roaops hybrid in the Marshall Islands. Photo by Raycrew Hiro.
In some of these hybrids with relatively clean nape regions, you can see where the idea of C. tinkeri occurring in Micronesia might have come from. However, as previously discussed, it is unlikely that the species (along with any other Hawaiian endemic) ever makes its way here.
Inter-subgeneric Hybrids
Hybrids involving other members outside of Roaops are rare, and curiously, only C. tinkeri has been documented to quench its libido with inter-subgeneric hybridization. In Hawaii, Chaetodon tinkeri hybridizes with C. miliaris and C. quadrimaculatus to form two exceptionally beautiful crosses. Despite being relatively rare, these hybrids have been documented with some regularity, suggesting that they may not entirely be stemmed from accidental fertilization. For example, C. tinkeri has been documented to form intentional pairs with C. tinkeri x C. miliaris hybrids. Seeing as all three species are common in Hawaii, why this unusual relationship occurs is hard to explain.

A hybrid of Chaetodon tinkeri x C. miliaris. The black back and dorsal spotting are clear traits of C. tinkeri. Photo by Richard Pyle and Robert Whitton.

Chaetodon quadrimaculatus x C. tinkeri. Both parental phenotypes are very evident here. Photo by Wayne Sugiyama.
It should be noted that inter-subgeneric hybrids are documented more often in butterflyfishes than other genera. C. capistratus x C. ocellatus, C. xanthurus x C. guentheri, C. lineolatus x C. auriga, C. daedalma x C. nippon, C. lunula x C. lineolatus, C. semilarvatus x C. lineolatus and C. multicinctus x C. miliaris are some examples of inter-subgeneric hybrids that are prevalent and well documented in butterflyfish. In many of these examples, both parents are sympatric, common, and not of the same clade, which would make hybridization rather unusual.

No other inter-subgeneric hybrids aside from C. tinkeri have been documented in the Roaops subgenus, but in all likelihood, this could just be due to poor documentation. C. burgessi should be expected to do the same, in part due to its wide ranging nature and sympatry with a large majority of other Chaetodon species.
In summary, the Roaops butterflies present us with a collection of highly intriguing findings. We have seen the extensive, yet aberrant distribution of this subgenus across the Indian and Pacific Oceans. We’ve also discussed the possible origin of the flavocoronatus phenotype, hypothesizing that it stemmed from the hybridization of at least two Roaops members, the combination of which differs in the Marianas and Micronesia. The hybrid hypothesis is supported by parallel examples seen in other reef fishes such as C. shepardi and C. katherinae.
This article should not be seen as a definitive answer with regards to the Roaops conundrum, but rather as an intuitive guide to the questions that need answering. With future studies and advanced genetics, perhaps the origin of C. flavocoronatus and how the other Roaops butterflyfishes relate will finally be elucidated.

0 Comments