


From the outset, the author aimed to provide awareness of the molecular mechanisms and exogenous cues that assist the remarkable events that precede and expedite procreative fruition in reef-forming corals, much of which remains an enigma. The previous editorials comprise essential background while a glossary in Appendix I clarifies some unfamiliar terms.
Numerous shallow-water zooxanthellate Scleractinia (hermatypes) spawn in synchrony once a year several days after the full moon (AFM) which implies this punctuality is coordinated by moon albedo or tidal-associated cues, whereas oo- and spermato-genesis can take anywhere from eight to 12 and from two to eight months in species of Acropora respectively (Prasetia et al. 2016).
It was discovered in the early 21st century that conspecific cohorts exploit recurrent broadcasting windows within mass coral spawns that deviate by merely seven and in some instances two minutes from year to year (Vize et al. 2005). This feat of uniformity is extraordinary insofar as corals do not have eyes or central nervous systems (CNSs; Brady et al. 2011). Yet these phenomena optimize the likelihood of intraspecific cross-fertilization whilst they hinder interspecific hybridization (Levitan et al. 2004; Baird et al. 2009) which more often than not, results in chromosome instability (Huret et al. 2000) and reproductive failure (Oliver & Babcock1992; Huret et al. 2000). All the same, scleractinian hybridization remains exceedingly rare (Bongaerts et al. 2021, cited in Furukawa et al. 2024; Ramirez-Portilla et al. 2022; Bridge et al. 2024; Furukawa et al. 2024).
Hundreds of millions of eggs are liberated by several hundred species (Babcock et al. 1986; Penland et al. 2004; Oldach et al. 2017) which optimizes the gene pool, satiates predators, and enhances procreative success (Jokiel et al. 1985; Lin et al. 2021).
Three types of gametocyte shedding were observed by Babcock and colleagues during the mass coral spawns on The Great Barrier Reef (GBR) in the spring of 1982, 1983, and 1984 (Babcock et al. 1986). Sperm and egg bundle packaging takes from 20 to 60 minutes before “setting” in the polyp pharynx where they rotate beneath the oral disc before liberation through the mouth (Type I). Conversely, brief “setting” was followed by cross anatomy polyp contraction and oral pore ejection. These convulsions were iterated minutes later in some which expedited further jettisoning (Type II). Moreover, gametes floated leisurely through the wide-open mouths of significantly aggressive Hynophora excesa (Type III; Babcock et al. 1986).
Most gamete bundles were positively buoyant and ascended to the surface where their cargo dispersed, whereas the eggs of Astreopora myriophthalma and Goniastrea favulus sank, but only after they had been separated from spermatozoa. Surface slicks likely enhance fertilization likelihood (Langley et al. 2024) where significant zygotic fusion was absent until two and a half hours after broadcasting, while the motility of planulae took around 36 hours to develop (Babcock et al. 1986). Some spermatozoa lose their viability after two hours while newly broadcast eggs resist fertilization (Levitan et al. 2004).
Annual cadences of average sea surface temperature (SST), monthly lunar and/or tidal cycles, and diel illumination are putative candidates that facilitate spawning synchrony insofar as mass broadcasting occurs on GBR from mid-October to December following spring upturns in SSTs from August to September. This deduction is sustained by the lunar month delay between inshore Magnetic and offshore Orpheus and Palm Islands where the former’s maximum 10 meter depth expedites warming, whilst other scleractinian spawns appeared attuned by mean SSTs (Kojis & Qninn 1981; Babcock et al. 1986; Baird et al. 2009). Solar daylength is central to spawning synchrony (Penland et al. 2004) while mean insolation appears not (Baird et al. 2009).
Most of GBR’s species of stony coral spawn AFM near to the next quarter moon whereas two out of over one hundred spawned after the inconspicuous new moon (●), whereas Acropora elseyi broadcast bundles after both. A rise in SST may assist gonad maturation inasmuch as corals with mature gametes spawned nocturnally after the next full moon (Babcock et al. 1986).
Artificial lunar cycles have prompted reproduction in Pocillopora damicornis (Jokiel et al. 1985) which suggests that circalunar periodicity may be entrained, while eminent observations in the late 20th century associated spawning with post-sunset darkness (Harrison et al. 1984; Babcock et al. 1986). Manipulations of the light/dark cycle in Goniastrea aspera determine the hour of spawning (Babcock 1984).
Harrison and collaborators were amongst the early pioneers to report mass synchronized spawns on GBR (Harrison et al. 1984) where low tide transpires from afternoon to midnight AFM with a minimum total displacement of merely half a meter, where most corals spawn in slack water. Goniastrea favulus routinely spawn on a low tide but during the day like Pocillopora verrucosa (Babcock et al. 1986; Bouwmeester et al. 2011); nevertheless, the procreative timekeeping of captive corals is induced by merely wildtype oscillations in light and darkness (Babcock 1984).
Most corals participating in GBR’s annual spring event do so on the third to sixth night AFM during the post-sunset/pre-moonrise period of darkness (Babcock et al. 1986; Lin et al. 2021). Broadcasting is spread over several months on different lunar days in the Red Sea (Shlesinger & Loya 1985) while the interspecific waterborne gametes of several Caribbean cohorts are barely separated like species of Acropora (Szmant-Froelich et al. 1984, cited in Babcock et al. 1986). However, neither of these instances are analogous to what occurs on GBR (Babcock et al. 1986). Variations in tidal amplitude and average SSTs appear to be more significant on Australia’s eastern coast (Maxwell 1968, cited in Babcock et al. 1986) compared to the Red Sea (Szmant-Froelich et al. 1984, cited in Babcock et al. 1986) and the Caribbean (Olhorst 1980, cited in Babcock et al. 1986), which conceivably entrains the spawning consortium’s endogenous noncircadian pacemaker (Babcock et al. 1986).

Fig 1. A map of coastal western Australia (WA) illustrating reef shelf composition where diverse milieus give rise to extraordinary communities. Map courtesy of Gilmour et al. 2016 and the Creative Commons Attribution License 4.0 (CC BY).
Brooded planulae or broadcast gametes are only disseminated up to 4 kilometers (Gilmour et al. 2016; Figueiredo et al. 2022), which assists their current poleward trajectory (Poloczanska et al. 2016). Brooders procreate over several months of the year, whereas egg and sperm broadcasters are typically constrained to one or two instances per annum (Gilmour et al. 2016).
Corals on western Australia’s Kimberley atolls tend to be gamete liberators where Acropora are limited to communal spring or autumnal spawns occurring in October/November or March/April with 10 and 90 percent respective participation. Diploastrea heliopora reproduced in spring whereas Favia stelligera and F. pallida procreated in both seasons. Western Australian (WA) surveys of oocyte coloration suggested that broadcasting took place on either the night of the full moon or on the next quarter neap tide seven to nine days later, but the day altered from year to year conceivably due to asynchrony between the lunar cycle and Gregorian calendar. Nevertheless, there is no period of darkness after sunset on the former (Lin et al. 2021). Massive Porites spawned moderately over several months from October to May on WA reefs, which differs from this genus inhabiting other Australian waters. Likewise, brooding Isopora brueggemanni and Stylophora hystrix also planulated each month from spring to autumn (Gilmour et al. 2016).
Spawning windows on shore-adjacent Kimberley reefs were analogous to oceanic, despite the majority of Acropora spawning in summer. There was nil evidence of the month earlier coastal spawns that occur on GBR compared to offshore (Gilmour et al. 2016). However, atoll formation is constrained on GBR because the continental shelf does not subside (Purdy & Winterer 2005) while corals adorning their slopes are not exposed to the tidal cues familiar to barrier or fringing reefs. Nevertheless, “split” spawning occurs in some species where corals broadcast on consecutive lunar months, and without doubt, several events remain undetected due to lengthy sampling interims and forecast bias. Autumn is likely when most inshore Kimberley corals reproduce with a minority split between April and March (Gilmour et al. 2016), whilst researchers remarked that inshore northern Indonesian corals exhibit protracted broadcasting over several hotter months with peaks occurring in spring and summer (Baird et al. 2009) which they could not rule out. The region’s non-acroporid brooding corals likely planulate over several warmer lunar cycles perhaps extending into winter (Gilmour et al. 2016).
The extensive near- and mid-shore reefs off western Australia’s Pilbara offer a variety of distinct environments with copious cross-shelf diversity like those of Montebello and Barrow Islands with most studies concentrated off the Dampier Archipelago. Historic data suggested split-spawning merely in autumn while later studies identified a minority of events occurring from October to December, and likewise, Favites flexuosa, F. pentagona, and Montipora undata likely spawn from spring to summer. Several of these reefs are dominated by massive Porites, Pavona decussata, and Turbinaria mesenterina which ostensibly undergo atypical hermaphroditic iteroparity culminating in a single spring or autumnal event (Gilmour et al. 2016; Orejas et al. 2023). However, later exhaustive histology discovered that these species were gonochoric whose trajectories varied. Pilbara’s Porites lobata spawn mostly in December (Gilmour et al. 2016) in harmony with their eastern Australian counterparts (Harriott 1983a; Harriott 1983), whilst Pavona decussata broadcast in both March and April possibly due to a WA split peculiar to 2007, whereas Turbinaria mesenterina likely procreate over several months from November to April. This region’s Porites lutea liberate gametes on spring tides from two to four days AFM in comparison to their conventional neap tide trajectory on the next quarter moon. The larval-rearing colonies of Pilbara likely planulate from spring to autumn and perchance early winter akin to Kimberley brooders (Gilmour et al. 2016).
The cooler waters off southerly Ningaloo are home to a 300-kilometre network of fringing reefs. Remarkably, mass autumnal spawns occur in March and April in the respective north and south which may reflect a split, with select broadcasting and planulation occurring over several months with merely conspecific effluxes in spring from September to October. 12 and 92 percent of Acropora liberated bundles in spring and autumn while one species shed in summer. 20 percent of the 69 widespread non-acroporans spawned in autumn with nil evident in spring. Most data were acquired in Coral Bay where Acroporidae and Faviidae participated in the mass autumnal event, while numerous species broadcast throughout summer and autumn typically on consecutive months. Legions of colonies spawned in unison late March to early April on a neap tide at Coral Bay seven to 10 days AFM, while a minority broadcast on the preceding or subsequent new or full moons. The April mass shedding that occurred AFM in late March 2002, has typically transpired a week earlier in other years. Less than 20 percent of the non-acroporid colonies participating in the early April event also either broadcast the first week in March or a minority in early May after the full or new moon. The comparatively tepid waters of Ningaloo influence gametogenesis which likely contributes to a procreative split between March and April. A. papillarae are known to spawn independently of their congeners with a summer trajectory in either December and/or January, whereas a limited number of Ningaloo’s colonies of hermaphroditic Echinopora lamellosa reproduce in February’s late summer whilst the majority procreate in autumnal April with a modicum in March (Gilmour et al. 2016).

Fig 2. Coral community composition on western Australian (WA) reefs. Analyses and chart courtesy of Gilmour et al. 2016 and the Creative Commons Attribution License 4.0 (CC BY).
The Houtman Abrolhos Islands lie near the geographic limit of tropical Scleractinia 100-kilometres off southwestern Australia. The genus Acropora and family Faviidae are the most widely studied, albeit investigations of broadcasting on other months remained scarce, in situ gametogenesis and aquarium spawning provided evidence for an autumnal meiocyte efflux in March. It is likely that the region’s patterns of procreative fruition are analogous to other WA communities, while egg and sperm liberation occurred on a low-amplitude tide from 10 to 11 and eight to 11 days after March’s full moon. Some studies provided evidence of April spawns with a putative three-week window of nocturnal broadcasting starting from a few days before and ending several days AFM. Studies of polyp gonads suggested that several colonies did not spawn that year or reproduced on other unexamined months, while planulation data were lacking (Gilmour et al. 2016).
Notwithstanding the current migration of corals to greater latitudes to escape the rigors of climate change (Poloczanska et al. 2016), few corals currently inhabit Australia’s southwest, where merely 26 species from 16 genera can survive at Rottnest Island. Liken that to near-shore Kimberley with 300 species of Scleractinia from 71 genera. Pocillopora damicornis monopolize temperate zones like the Mediterranean (Hunter 1993; Ayre & Miller 2004; Miller & Ayre 2004) where they may brood asexual planulae (Ayre & Miller 2004; Miller & Ayre 2004; Schmidt-Roach et al. 2012). Conversely, clonal replicative strategies such as fragmentation (Crane 1999, cited in Gilmour et al. 2016) or bail-out where a settling polyp is ejected from the colony (Sammarco 1982; Capel et al. 2014) may assist establishment of Acropora and Porites lutea in temperate zones. Alveopora fenestrata, Montipora mollis, Pocillopora damicornis, Symphillia wilsoni, and Faviidae family affiliates predominate regionally with species like Goniastrea aspera and G. australiensis that favor cooler waters. Nevertheless, fleshy macroalgal cover is ~60 percent, while the diminished-diversity reef at Hall Bank boasts around 50 percent Scleractinia comprising 14 species where the genera Favites and Goniastrea prevail (Thomson & Frisch 2010; Gilmour et al. 2016). Coral spawning and planulation had only been studied off Rottnest Island which inferred summer through autumn from December to April commensurate with reproduction on GBR (Gilmour et al. 2016), which is extraordinary inasmuch as SSTs are known to influence gametogenesis and the synchrony of broadcasting (Kojis & Qninn 1981; Babcock et al. 1986; Keith et al. 2016). Regional P. damicornis broadcast both kinds of meiocytes (Ayre & Miller 2004; Miller & Ayre 2004) and brood (Schmidt-Roach et al. 2012; Gilmour et al. 2016), while the spawning of Alveopora fenestsrata and Symphyllia wilsoni appears circa-attuned to the inconspicuous new moon (Gilmour et al. 2016) which may reflect the response to darkness or diminished lunar reflectance documented by Babcock and researchers in 1986 and Lin and collaborators in 2021. Temperate water appears to expedite a protracted reproductive season that extends over two months for prevalent corals like P. damicornis and Turbinaria mesenterina which breed in summer and autumn respectively. The researchers proposed that lower southerly SSTs may hinder the entrainment of precise spawning windows in all but the height of summer. Tropical corals exploit divergent reproductive strategies in cooler water where Plesiastrea versipora is inordinately abundant in south Australia while its ecology remains enigmatic (Gilmour et al. 2016).

Fig 3. The percent reproductive output of corals from spring to autumn on various western Australian (WA) reefs. Analyses and graphs courtesy of Gilmour et al. 2016 and the Creative Commons Attribution License 4.0 (CC BY).
“Keystone” corals upon which much of an ecosystem relies may constitute a mere minority. Field studies identifying and elucidating their natural history are required to constrain local anthropogenic agendas to months which minimize disruptions of their gametogenesis and spawning. In situ studies exploit fragmentation where damaging rarer taxa may prove environmentally unsound, yet specimens can be semi-permanently tagged and sampled in rotation. All the same, accounts of winter spawnings are rare, so activities such as dredging and ballast acquisition can be focused offseason (Gilmour et al. 2016).
Remarkably, corals inhabiting greater latitudes with extended daylight spawn closer to the full moon, while inshore underwater anthropogenic light pollution has an analogous effect (Fig 4.; Davies et al. 2023); hence near-shore corals cannot cross-fertilize with their mid-shelf or offshore counterparts, which restricts the gene pool possibly weakening species integrity. Nevertheless, corals exploit epigenetic modification (Gittins et al. 2015) and are significantly phenotypically plastic (Million et al. 2022), whereas gametes, and even planulae, are not dispersed vast distances (Figueiredo et al. 2022). Investigations of broadcasting and planulation are best carried out in aquaria because species are challenging to discern in situ, thus many studies differentiate colonies merely to the rank of genus or family, while invasive pre-spawning collections can perturb the most fecund (Gilmour et al. 2016).
Modern dimmable, software-controlled ballasts that emulate natural cycles of solar and lunar irradiance are readily available. However, the most versatile manipulate light emitting diodes (LEDs) which prove disastrous for the long-term culture of small polyp hermatypes, inasmuch as their emissions are noncohesive and lack essential up- and side-welling (backscatter). Hence successful husbandry relies on fluorescent tubes and metal halide high intensity discharge (HID) lamps. Moreover, most researchers are unfamiliar with the intricacies of captive culture and as such, are unable to accurately emulate pristine wild milieus which confounds objective science. For instance, if propagules in the control groups are bleaching or losing flesh in ostensibly survivable conditions, no matter how negligeable, then it is more than likely that they are being exposed to at least one unconsidered confounding biochemical process. Corals do not lose pigment or flesh when they are supplied with everything that they need, and nothing that they do not need. Consult: The Complete Reef Aquarist for further guidance (https://www.ebay.co.uk/itm/116390701502).

Fig 4. Histograms of lunar days where x = 0 equates to a full moon while spawning frequency is plotted on the y axis for corals in the East China Sea; on The Great Barrier Reef; in the Gulf of Thailand; the Persian Gulf; the Red Sea; the Gulf of Eilat; the South China Sea, and the Sulu, Celebes, Banda, and Java Seas. Overhead represent mean spawning windows AFM for corals that were exposed to artificial light at night (ALAN; pink) and those that were not (sky blue). Most ALAN-exposed corals spawned between one and three days earlier, which is a likely consequence of nominal underwater illumination during the period of darkness between sunset and moonrise on evenings after the night of the full moon (Lin et al. 2021; Davies et al. 2023). Spring broadcasting occurs later in the season in corals furthest from the equator (Keith et al. 2016). All things considered, such findings are redolent of sunset adjusting the procreative pacemaker (Penland et al. 2004). Analyses and histograms courtesy of Davies et al. 2023 and the Creative Commons Attribution License 4.0.
Palau’s most momentous spawning event of 2002 occurred in the autumn, while less significant mass broadcasting transpired in late May four to five days AFM, which was repeated in spring six to seven days AFM in late August, and again in late September one to three days AFM. Another multispecies spawn occurred during the height of summer two days AFM in February 2003 (Tables 1. & 2.; Fig 7.; Penland et al. 2004).
Summer spawns are extraordinary insofar as corals must expend significant resources on constituent immunity and surplus excitation-dissipating protective strategies that are not required in winter (Toledo-Hernandez et al. 2023) which may be why Palauan autumnal spawns are the most profound, when insolation maxima subside (Penland et al. 2004).

Table 1. The 2002 autumnal spawns of several of Palau’s Scleractinia. Adapted from Penland et al. 2004.
Broadcasting appeared uninfluenced by depth on Indonesian reefs while autumnal events coincided with elevated SSTs in May 2002, but with diminished temperatures in September, October, and February when several species of Montipora spawned on multiple occasions in spring and summer (Table 2.; Fig 7.; Penland et al. 2004). Keith and allies recognized that sharp rises in SSTs and moderate wind appeared to determine the month of spawning in corals inhabiting the Indo-Pacific, while the impact of average temperatures remained unclear. Modest breeze augments the horizontal advection and dissemination of gametes (Keith et al. 2016).
The southern oscillations of El Niño and La Niña further impact global as well as Indonesian weather systems where the former’s high pressure and diminished clouds spike sunlight over Palau in August (Figs 5. & 6.; Penland et al. 2004).
Tides are elicited by the gravity of the sun and moon (Guest et al. 2002; Penland et al. 2004) while insolation maxima occur twice a year on the vernal and autumnal equinoxes around the 20th or 21st of March and the 22nd or 23rd of September when the sun crosses the equator which confers a 12-hour day (Ruggles 1997; Penland et al. 2004; Sager & Mc Farlane 2017). Celestial body alignments around the full or new moon create exaggerated “spring” tides (Ruggles 1997) which likely prompt anomalies in SSTs that modify reproductive chronobiology. The rise and fall of insolation maxima correspond to the mass spring and autumnal spawns in Palau (Penland et al. 2004).
The current study discovered that the vernal and autumnal peaks in solar irradiance coincided with the mass broadcasting events on GBR and in Japanese waters, where corals on southern and northern reefs spawned in spring (May) and in summer (July/August), whereas such extremes of photon flux density range from 5,500 W m-2 d-1 (~11,550 µmol m-2 d-1) in Palau to 6,550 W m-2 d-1 (~13,655 µmol m-2 d-1) in Australia and Japan (Penland et al. 2004). An analogous three-month disparity between GBR’s central to southern coral communities was evident while Keith and collaborators recognized that the spawning seasons were separated by a month in the northern and southern hemisphere (Keith et al. 2016). Corals on reefs 19o south of the equator broadcast in spring around October/November whilst those 25o south procreate in November/December. Conversely, reproduction of colonies 30o south was delayed until summer from January to March.
Sunlight irradiating the earth has a direct impact on sea surface temperature, yet there is a lag between photons received and oceanic warming (Penland et al. 2004). Montastraea annularis spawned in Jamaica when average SSTs were highest in August just before the rainy season, where researchers postulated broadcasting was related to rainfall and temperature (Mendes & Woodley 2002). Stenohaline sessile invertebrates like corals and sponges may detect changes in salinity which could drive the reproductive pacemaker. Certainly, deluge-derived hyposalinity and terrestrial eutrophicants destabilize coral physiology and ecology. The study proposed that the peaks in insolation observed in January/February may be an exogenous cue that initiates gametogenesis eight months before the fruition of procreation in August/September, when the autumnal equinox’s solar irradiance could instigate further gonad development with a broadcasting trajectory in the following April/May (Fig 7.). Hence three cycles of gametogenesis would be completed every two years, whilst corals at greater latitudes spawn merely once per annum (Penland et al. 2004).
Remarkably, Nikko Bay’s elevated turbidity curtails downwelling which assists coral survivorship in the face of ferocious solar irradiance and heat. The study concluded that cycles of insolation coincide with scleractinian reproduction across the western Pacific Rim (Penland et al. 2004).
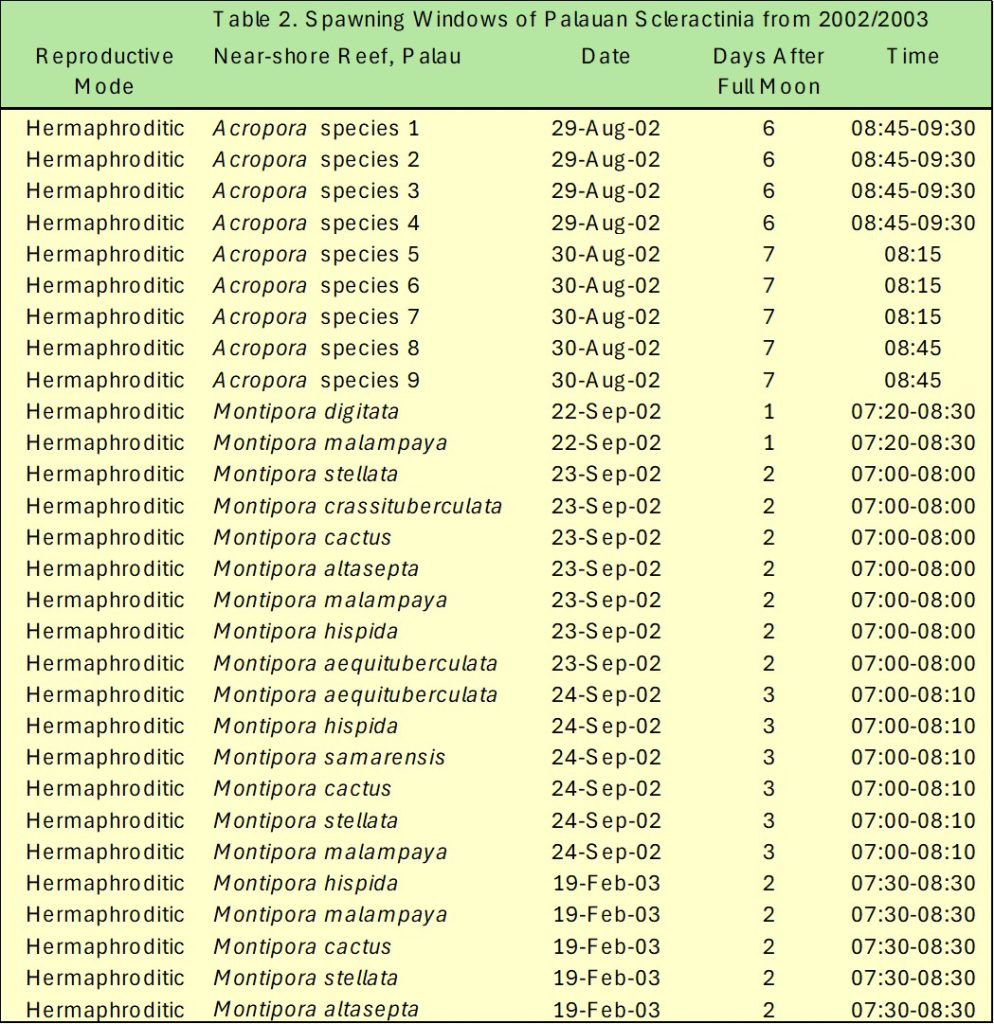
Table 2. The 2002 to 2003 spring and summer spawns of several Palauan Scleractinia. Adapted from Penland et al. 2004.
Brood colonies of P. damicornis planulate every lunar cycle with a full moon peak in spring and another less meaningful efflux around the new moon in summer. The sun’s ultraviolet (UV) emissions appear cyclic while its rotation takes 27 days (Jokiel et al. 1985). Intensities of UV impact larval profusion in this species (Jakie & York 1982) while moonlight remains proportional to solar radiation. Cycles associated with tidal ebb and flow such as jostling, pressure, and temperature may act as rhythm-entraining cues (Jones & Naylor 1970; Hastings 1981; Zhang et al. 2013). Detection of lunar albedo or the consequential abundance of zooplankton are candidate zeitgebers which may attune the endogenous noncircadian reproductive pacemaker. However, geographically-distant colonies release larvae at different times. P. damicornis exploit various proliferative strategies whereas merely their brooding and sperm broadcasting were considered, where peaks of planulation in type B colonies consistently occurred between the first quarter and full moon, whereas larval shedding spiked in type Y corals around the third quarter moon (Jokiel et al. 1985).
Genetically distinct Hawaiian P. damicornis types Y and B planulate around the full or new moon while the former retains a yellow hue. Overnight disruption and perturbation of the periodicity of spawning had been observed during cloud cover and continuous dim light (Stimson, personal communication, cited in Jokiel et al. 1985). Some of Jokiel and colleagues’ pioneering aquarium-based experiments ascertained whether corals exposed to various lighting protocols would planulate in synchrony with their wild cohorts. Their study aimed to highlight the light-responsive nature of coral reproduction or unearth the existence of an endogenous procreative “clock”. Groups of colonies were exposed to daytime natural light from 05:00 to 19:00 followed by a continuous full moon-analogous PAR of 0.01 µmol m-2 s-1, or new-moon-comparable continuous darkness over an entire month, or an artificial phase-shifted lunar cycle with utter darkness on the night of the full moon but with 10 days of simulated moonlight centered around the new moon (Jokiel et al. 1985).

Fig 5. The El Niño southern oscillation (ENSO) where cooler water in Peru is ordinarily derived from the upwelling of nutrient-rich deepwater.
Emulations of candidate non-photonic circatidal-entraining zeitgebers did not impact spawning synchrony in P. damicornis but highlighted the stenohalinicity of this species and its predisposition to exploit parthenogenesis to brood asexual planulae within two weeks from mature ova (Jokiel et al. 1985).
Cyphastrea ocellina and Tubastraea coccinea do not planulate monthly, while Hawaiian Fungia scutaria and Montipora broadcast gametes shortly AFM. The outcomes of experiments affirmed that planulation was synchronized by moonlight in P. damicornis insofar as nocturnal phase shifts led to entrainment of procreative periodicity, while utter darkness or continuous full moonlight diminished reproductive output. Remarkably, the larval liberative synchrony of type Y colonies was sustained for two months before aligning with the new lighting protocol, whereas type B corals retained merely one cycle before acclimation. This strain fails to planulate during times when moonlight fluctuates which is common during heavy cloud cover and volcanic activity, whilst previous evidence suggested that oogenesis took five weeks in type Y (Martin-Chavez, cited in Jokiel et al. 1985) which may explain this variant’s two-month reproductive inertia. Far more larvae were liberated from corals exposed to artificially phase-shifted moonlight in comparison to the moderate numbers shed by corals in dim light or the fewer still brooded and released by colonies exposed to unwavering diel darkness. However, significantly fewer spats settled/survived in the group exposed to a nighttime simulated continuous full moon (Table 3.; Jokiel et al. 1985).
P. damicornis’ three-day-old first polyps with mouths, tentacles, and mesenteries can abandon their nascent skeletons and reverse metamorphose into tentacle and mouth-bearing zooplankton (Richmond 1985). Months of continuous spat settlement and reverted aposymbiotic planulae would require smaller zooplanktonic prey to survive, which may explain the reduced numbers of glass-adhered juveniles. The groups of colonies exposed to various lighting protocols were housed with omnivorous fish and juvenile propagules were not enumerated until the conclusion of each experiment. Fish voraciously consume eggs and larvae (Westneat & Resing 1988; Pratchett et al. 2001; Lin et al. 2021) which may clarify why corals mostly broadcast and planulate at night. Fish will not feed in profound darkness, yet ambient irradiance stimulates their appetite (Wilson, personal communication; Aslett 2024) hence zooplanktonic planulae may be eaten throughout simulated full moonlight (Jokiel et al. 1985). However, the daytime-liberated gametes of P. verrocosa appear to be unpalatable (Bouwmeester et al. 2011).
The researchers proposed cross-anatomy polyp fecundity might be regulated by a light receptor or host-derived molecular signal because coordinating the reproductive status of each polyp would require “cross-talk” or the detection of an exogenous cue. The study concluded that planulation is light- and possibly temperature-responsive in P. damicornis (Jokiel et al. 1985).

Fig 6. The La Niño southern oscillation (LNSO) where cooler water is derived from nutrient-rich upwelling where the descent of dry air spikes insolation over Peru (Fig 5.).
SSTs appear to play a central role in gametogenesis which in turn can determine the month/day of spawning (Lin & Nozawa 2023) where corals on warmer reefs broadcast earlier. Acropora humilis spawn later than other Red Sea acroporids, where weather alters from month to month which in turn impacts larval distribution and thus community composition and restoration of coral cover. Numerous species of Acropora at respective Thuwal and Shushah liberate gamete packets in April and May on the night of the full moon (Osman et al. 2024) which may indicate that post-sunset darkness is not required for this genus (Lin et al. 2021; Osman et al. 2024). Lunar reflectance attunes the transcription of several candidate circadian genes implicated in the procreative fruition of A. millepora, which may determine the night of broadcasting (Brady et al. 2016).
A. humilis, Porites lutea, and P. solida do not spawn on the evening of the full moon (Hanafy et al. 2010, cited in Davies et al. 2023; Bouwmeester et al. 2014, cited in Osman et al. 2024; Attalla 2016, cited in Davies et al. 2023), when reproduction may be deferred by SSTs (Osman et al. 2024). Extended seasonal daylengths common to greater latitudes cause corals to spawn closer (Davies et al. 2023) while Dipsastraea speciosa’s is triggered by the period of darkness that follows sunset on evenings after the night of the full moon (Babcock et al. 1986; Lin et al. 2021). Nevertheless, rapid shifts in SSTs have caused western shaw Red Sea corals at Hurghada to spawn a few days beforehand. Osman and colleagues’ data strongly suggest seawater warming leading up to the night of spawning determines the broadcasting trajectory of Red Sea corals, yet the researchers remarked that other hitherto unidentified environmental signals influence spawning windows (Osman et al. 2024). Penland and collaborators found that elevated mean SSTs were associated with autumnal spawning off Palau, whilst cooler waters appeared to coincide with broadcasting in later months (Fig 7.; Penland et al. 2004). Wind abatement above tranquil seas (van Woesik 2010), the availability of food (Fadlallah 1983, cited in Osman et al. 2024), the spectral components of twilight (Boch et al. 2011), and rainfall (Mendes & Woodley 2002) may influence spawning synchrony (Keith et al. 2016; Osman et al. 2024).
The findings of Brady and allies published in 2009 suggested that sunset determined the time of spawning; however, a one-hour shift in simulated moonrise instigated an equivalent alteration in the spawning period, so even core entrained rhythmicity may instantaneously adapt to minor variances of up to an hour (Brady et al. 2009, cited in Brady et al. 2016; Brady et al. 2016). Nevertheless, their study indicated that the candidate circadian genes: Cry1; Cry2; Clock; Cycle, and Timeless are not under the control of an endogenous circalunar oscillator (Brady et al. 2016). Levy and colleagues (2007) documented elevated transcription of A. millepora’s Cry2 at midnight on the night of the full moon compared with the new moon, but unlike the current study, failed to detect a transcriptional increase in Cry1. The subsequent findings of a related team discovered that the expression of Cry1 was indeed raised on the night of the full moon and more so than Cry2 (Kaniewska et al. 2015), whereas Brady and allies detected a greater abundance of Cry1’s mRNA on the night of the new moon with a peak on the first quarter moon (Brady et al. 2016).
Cry2 appears significantly circadian in A. millepora and Nematostella vectensis insofar as this gene’s ~24-hour transcriptional rhythmicity was upheld for around three days in utter darkness (Peres et al. 2014; Brady et al. 2016). However, profusions of mRNA lacked circalunar rhythmicity over extended periods of darkness or full moon luminescence in all candidate circadian genes excluding eyes absent (Eya), in continuous full moon–exposed A. millepora. Nevertheless, the study verified that light alone regulates the spawning pacemaker of this coral like it does in others. Therefore, anthropogenic light pollution or climate change-driven cloud cover (Brady et al. 2016), has the potential to reduce the gene pool by manipulating the spawning windows of discrete regional subpopulations.

Fig 7. The relationship between sea surface temperature (SST), solar irradiance (insolation), and the timing of the mass spawning events in Palau from 2002 and 2003. Adapted from Patrick Colin’s data presented in Penland et al. 2004.
The findings of most studies suggest lunar cycles “trigger” gametic broadcasting but not all corals spawn nocturnally (Baird et al. 2009). Although undoubtedly some species broadcast meiocytes on the night of the full moon and reproductive cycles have a periodicity of ~30 days, the majority do not broadcast or planulate on a spring tide, full or new moon (Linden et al. 2018). Furthermore, typical tidal cues do not seem to impact spawning synchrony (Jokiel et al. 1985). Linden and collaborators suggested that scleractinian procreation was circatrigintan which describes any cycle with a periodicity of approximately 30 days that is not associated with exogenous cues.
Stylophora pistillata is conspicuous amongst the corals whose procreative curve appears independent of moonlight (Fig 8.; Rinkevich & Loya 1979; Rinkevich & Loya 1987; Amar et al. 2007). Merely two studies have associated S. pistillata’s spawning with lunar cyclicity (Fan et al. 2002; Zakai et al. 2006), whilst colonies of this gonochoric brooder appear to “trickle” planulate daily over several months from winter in December/January to mid- to late-summer in July/September (Linden et al. 2018).
The study highlighted the diverse iterated peaks of reproductive output amongst five of the sampled colonies whose periodicity ranged from 25 to 35 days, while planulation failed to spike in three. Polyp fecundity approached 100 percent from April to June during which the abundance of planulae escalated from corals 1 and 2, whereas those from the remaining six diminished on consecutive months (Tables 4. & 5.). Despite compelling evidence to the contrary (Tanner et al. 1996; Mercier et al. 2011; Brooke & Järnegren 2013; Keith et al. 2016), daytime solar and/or ultraviolet (UV) radiation and SSTs had a negligible impact on larval liberation (Linden et al. 2018). However, cross-correlation analyses identified a circa-lunar and -tidal rhythm to the peak planulation trends of five colonies where periodicity ranged from 27 to 33 days with a delay of five to 10 days AFM. Shedding was loosely associated with moonlight insofar as it transpired before or AFM and on a spring or neap tide (Fig 9.; Linden et al. 2018).

Table 3. Planula enumerations from types Y and B of P. damicornis exposed to simulated continuous full moonlight, new moon utter darkness, and nil illumination on the night of the natural full moon with 10 days of subjective moonlight around the time of the authentic new moon. Features the numbers of juvenile colonies/first polyps growing on randomly selected 8- by 20-centimeter strips of the aquarium glass after each experiment. Adapted from data presented by Jokiel et al. 1985.
Rinkevich and Loya reported declinations in S. pistillata’s planulation throughout their experimental periods from 1979 to 1987 consistent with the current study. The output of three colonies appeared to lack a procreative pacemaker, while the peak progeny efflux of two of the six that remained, exhibited clear periodicity between 25 to 27 days. Even so, the patterns of another three corals ranged from 33 to 35 (Linden et al. 2018).

Fig 8. Various in situ morphotypes of Stylophora pistillata in Eilat’s Red Sea whose reproductive patterns do not appear to conform to phases of the lunar cycle. Plate “h” illustrates the yellow growing tips of type Y, while the scale bar of plate “a” represents 3 centimeters (Linden et al. 2018). Images courtesy of Linden et al. 2018 and the Creative commons Attribution License 4.0.

Table 4. The mean totals and maximum planulae shed from colonies of Stylophora pistillata per day between April and June off Eilat in the Red Sea where colony size is positively correlated with planulation volume (Linden et al. 2018). Table, analyses, and surveys courtesy of Linden et al. 2018 and the Creative Commons Attribution License 4.0.

Table 5. The mean number of planulae shed from colonies of Stylophora pistillata per day in April, May, and June off Eilat in the Red Sea amongst which the reproductive output of the largest colony remains conspicuous (Table 4.; Linden et al. 2018). Table and analyses courtesy of Linden et al. 2018 and the Creative Commons Attribution License 4.0.

Fig 9. A graph illustrating the relationship between the new and full moons of April to June and solar radiation, sea surface temperature (SST), tidal amplitude, and peak planulation in colonies of Stylophora pistillata in the Red Sea. Graph and analyses courtesy of Linden et al. 2018 and the Creative Commons Attribution License 4.0.
The experimenters remarked that despite their rigorous observations and trapping protocols, they could not rule out unobserved larval shedding (Linden et al. 2018). Even though the cross-correlation analyses unearthed a circalunar and circatidal periodicity to planulation in some colonies, none had cyclicities close to 29.5 days, and thus in the absence of adaptive mechanisms, would drift in and out of phase with spring and neap tides and full, new, and quarter moons (Fig 9.; Linden et al. 2018). Such findings lend credence to the zeitgeber-independent circatrigintan paradigm (Linden et al. 2018).
Cross-fertilization may be tightly shackled to the lunar cycle in S. pistillata which has only been surveyed in a couple of studies (Rinkevich & Loya 1979a; Rinkevich & Loya 1979) while those evaluating the regularity of sperm efflux are lacking. Strict circalunar spermatogenesis and cross-fertilization may still give rise to unsynchronized planulation if embryogenesis is shorter or longer than 29.5 days, which takes two weeks in P. damicornis (Jokiel et al. 1985; Linden et al. 2018).
Eilat’s significant artificial light at night (ALAN; Davies et al. 2023) no doubt impacts and perhaps decouples the larval efflux pacemaker, which would clarify why some corals did not exhibit stringent timekeeping, whereas the peak reproductive output of others was coordinated and cyclic (Linden et al. 2018).

Fig 10. Illustrating the proposed underlying means by which some of Acropora millepora’s daily transcripts could align with key events in a lunar cycle. Top – transcriptional amplitude and frequency remain constant but expression moves to accommodate peaks and troughs on a different kind of moon and time of day. Bottom – expression remains in phase but the profusion of mRNA alters. Arrows indicate midnight on a new or full moon. Adapted from Brady et al. 2016.
The transcription of gene sets associated with mitosis and cell cycle exhibited a robust circalunar rhythmicity in species of Acropora peaking on the night of the full moon (Oldach et al. 2017) while the reproduction of several marine teleosts appears attuned to moonlight (Fukushiro et al. 2011).
By virtue of planetary spin, the same positions on the earth and moon realign every 24.8 hours, which elicits two high and low tides with 12.4-hour periodicities. The moon takes 29.5 days to orbit the earth which in conjunction with solar reflectance, forms the familiar lunar cycle. The Gregorian calendar, Greenwich meantime (GMT), and lunar months thus drift in and out of phase yet synchronize at the same time of day every 14.8 days on a spring or neap tide (Bünning & Müller 1961; Naylor 1996; Neumann 2014; Kaiser & Neumann 2021; Rock et al. 2022). The current and a recent study observed that 24-hour gene expression either shifted phase to align with major phases in a lunar month whilst retaining periodicity and amplitude, or transcription remained aligned while mRNA abundance changed (Fig 10.; Fig 11.). These models and the research findings which support them, suggest unlike cueing, the phases of the moon modulate scleractinian expression, which impacts the endogenous circadian oscillator (Brady et al. 2016; Oldach et al. 2017).

Fig 11. Exemplifying the phase shift of a model gene’s transcriptional peaks and troughs to align with a lunar cycle’s key events consistent with the observations of Brady and collaborators from 2016 and Oldach and colleagues from 2017 (Fig 10.). Courtesy of Oldach et al. 2017 and the Creative Commons Attribution License.

Fig 12. The routine spawning of the prevalent corals inhabiting the Gulf of Mexico’s Flower Garden Banks (FGB) National Marine Sanctuary on the evening of the eighth day AFM from August 1997 to 2003, where dark grey boxes represent core broadcasting, while window peripheries are specified by light grey. Pseudodiploria strigosa’s (synonym: Diploria strigosa’s) gradient plot illustrates efflux with a sizeable early bias. Of note are the timekeeping disparities between spermatozoan and oocyte liberation from gonochoric Montastraea cavernosa and Stephanocoenia intersepta. Adapted from data presented by Vize et al. in 2005.
Conspecific broadcasting within the annual Augustan spawn is unique inasmuch as excluding Pseudodiploria strigosa, no other corals liberate their gametes at the same time as Colpophyllia natans, Montastraea annularis, M. cavernosa, M. faveolata, M. franksi, and Stephanocoenia intersepta in The Gulf of Mexico’s Flower Garden Banks (FGB) National Marine Sanctuary. Nevertheless, euphotic and mesophotic dwelling corals sharing the same geographic location spawn in synchrony irrespective of irradiance downwelling and SSTs. It is thought that mean SSTs determine the month, lunar periodicity sets the day, and sunset prompts the hour (Fig 12.; Vize et al. 2005). The broadcasting of spermatozoa occurs weeks before planulation.

Fig 13. Illustrating day- and night-time wildtype illumination leading up to and following the full moon, which triggers spawning in Dipsastraea speciosa from 21:00 to 22:00 on the 6th evening AFM, because spawning is triggered by the post-sunset darkness on the night of day 1. Adapted from the Lin et al. 2021.
The year to year procreative windows of Colpophyllia natans are exceedingly consistent insofar as they commence within 2 minutes and end within 5 in The Gulf of Mexico. Conversely, the timekeeping of Pseudodiploria strigosa is the least precise. Gonochoric Montastraea cavernosa and Stephanocoenia intersepta liberate spermatozoa before ova. Paternal M. cavernosa shed gametes five to 23 minutes before female colonies which appeared independent of nearby males, whereas maternal Stephanocoenia intersepta do not liberate eggs unless a male releases sperm within half a metre. Most eggs of this species were fertilized before collection which may suggest fertilization “triggers” their distribution (Vize et al. 2005).
The spawning windows on FGB reefs are largely analogous to those of the Caribbean but less protracted. Nevertheless, the onset of Panamanian and Bahamian M. faveolata’s broadcasting is delayed by 30 to 40 minutes and ends 40 to 60 minutes later than their remote regional counterparts (Vize et al. 2005). Levitan and collaborators reported that M. faveolata spawned later than M. annularis in 2004 which is reversed at FGB. M. franksi’s broadcasting occurred 20 minutes later off Panama and the Bahamas, while such evidence implies that corals retain local environmental adaptation competencies (Vize et al. 2005).
Sunset ostensibly determines the hour of spawning (Brady et al. 2009, cited in Lin et al. 2021), where Orbicella franksi and O. annularis spawn two and four hours after sunset (Levitan et al. 2004), whilst moonlight appears to determine the day (Lin et al. 2021). The prevailing paradigm was broadcasting was expedited through cryptochrome detection of lunar reflectance, downstream signal transduction, activation of transcription factors, and modified gene expression (Levy et al. 2007; Kaniewska et al. 2015).
The seminal study of Lin and colleagues devised a series of simple light blocking and dim illumination protocols to prove that spawning is “triggered” by the period of darkness between sunset and moonrise on evenings after the night of the full moon, whereas the moon ascends before sunset on nights leading up to and on the evening of the full moon (Figs 13. & 14.). When colonies of Dipsastraea speciosa were covered during nights around the full moon, they routinely spawned on the fifth night after the onset of darkness. Dim light at night also delayed broadcasting irrespective of wavelength that occurred five and six days after colonies were shaded. Spawning was delayed by four nights in corals exposed to three to four days of dim irradiance which appeared to impact synchrony, while there was no delay in corals exposed to subdued luminescence on the latter half of subjective night. Spawning from 21:00 to 22:00 remained consistent in the wild and in corals under all experimental conditions whereas one hour of post-sunset darkness was sufficient, but the competency of gametic release was elevated in corals obscured for the entire night. These mechanisms were unreliant on short wavelength blue which suggests that cryptochromes and peripheral cells expressing r-opsin are not used to detect moonlight (Shoguchi et al. 2013; Lin et al. 2021; Revilla-I-Domingo et al. 2021).
That concludes our exploration of coral biorhythms which influence all aspects of scleractinian natural history. Science is proceeding at breakneck speed where new discoveries and open-access literature will deliver the means to slow the current decline in wild reef ecology.

Fig 14. Lin and collaborators recognized that moonrise occurs before sunset on evenings leading up to and on the night of the full moon, before which uninterrupted illumination suppresses spawning; however, the post-sunset darkness on the evening after the night of the full moon, lifts suppression which sets in motion molecular mechanisms that culminate in the broadcasting of Dipsastraea speciosa five nights later (Fig 13.). Adapted from Lin et al. 2021.
Appendix 1 Glossary
Allele refers to different forms of a gene.
Amplicon is an amplified sequence of DNA typically in a solution of its countless replicates (Merriam Webster 2018).
Animal Pole is a surface zone of an egg which marks the most mitotically-active yolk-diminished region.
The class Anthozoa comprises the polyp morphotypes corals, anemones, sea fans, and sea pens, whose non-mineralized ancestors emerged five hundred and fifty million years ago.
Aposymbiosis – when symbiotic organisms live separately.
Archenteron is the rudiments of a developing gut.
Autotrophic organisms satisfy their nutritional requirements from simple inorganic chemicals and usually but not exclusively light.
Azooxanthellate is an organism without symbiotic “algal”-partners.
Bathymetric refers to the environmental dynamics of deep and diverse ecosystems which vary in temperature, salinity, dissolved gasses, current, and pressure.
The infrakingdom Bilateria is a clade of bilaterally symmetrical animals with anteriors, posteriors, dorsums, flanks, and ventrums like dogs, humas, flukes, and mice.
Blastomere is a discrete nucleated cell formed from the mitotic division of a fertilized egg’s cytoplasm.
Blastopore is an opening that develops from an ectodermal invagination during the gastrulation phase of a developing embryo which usually becomes the mouth.
Bootstrap replicates are equal-sized multiple sequence-aligned fictional substitute nucleic acids with confidences proportional to percentage identity.
Canonical, substantiated; acknowledged.
Chorion is an embryo-enveloping membrane.
Clade describes a class of organisms comprising all the descendants from a common ancestor.
Codons are triplet bases in mRNA that encode for one kind of Amino Acid.
Conspecific means the same species.
Contigs are DNA molecules made up of smaller overlapping Oligonucleotides.
The cells of a Diploid organism have a chromosome from each parent (2n), whereas the copy number of Haploid organisms is 1n.
The suffix “cyte” means cell.
Diurnal simply means daily or daytime, yet contextually it refers to a diel cycle of typical daylight followed by nighttime-analogous darkness.
Endocrine refers to hormone-secreting ductless suprarenal, thymus, thyroid, parathyroid, pituitary, pancreas, ovary, and testis glands.
Endogenous factors originate from within.
Endonuclease, see Nuclease.
Epiboly refers to the thinning and spreading of a multiple layer of cells during gastrulation.
Exon is a gene integral coding sequence separated by an Intron (Dale 1998).
Expression, see mRNA.
Genets are culture-ready sexual or vegetative coral larvae (Planulae).
Germ refers to a cellular lineage or layer derived from one kind of embryonic Stem Cell or Progenitor. Anthozoa are Diploblastic because their ecto- and endo-derms develop from two discrete progenitors.
Gonochoric organisms are female or male.
Hermatypic and Ahermatypic Corals – the terms hermatypic and ahermatypic have been misused. Irregularities such as sea fans possessing rigid skeletons are constructive, but they may or may not contain “algal” symbionts and do not contribute to reef substratum. Ahermatypes may have large or small polyps and do not contain zooxanthellae. Scleractinia are “true” reef builders but may be azooxanthellate or contain “algal”-partners. Scleractinia and hermatype are not interchangeable. Hermatypic corals contain zooxanthellae and are constructive. Ahermatypes are azooxanthellate and non-constructive. This clear definition also applies to non-Scleractinia (Schuhmacher & Zibrowius 1985).
Heteropole refers to an imaginary core axis that spans the anterior oral and posterior aboral ends of an organism.
Heterotrophs cannot utilize inorganic carbon for growth so they must acquire organic plant- or animal-derived nourishment. Their sources include glucose, yet several microscopic taxa retain the capacity to transform inorganic compounds which are frequently used as substrates for chemical reactions.
Heterozygous refers to different forms of a gene usually originating from each parent on opposing chromosomes in a diploid (2n) organism where each one of an individual’s 1n gametes carry one kind of allele.
Holoblastic Cleavage utterly divides a fertilized egg’s blastomeres.
Homozygous refers to the same form of a gene usually originating from each parent on opposing chromosomes in a diploid (2n) organism where each one of an individual’s 1n gametes carry the same kind of allele.
Intron is a gene-integral interrupting (intervening) sequence that is excised during splicing (Dale 1998).
The yolk of Isolecithal ova is dense, granular, and extensively distributed.
Macronemes are thickened mesenteries that develop gonads in the starlet sea anemone, Nematostella vectensis.
Meiocyte is a cell generated by meiosis which is contextually synonymous with gamete, within which Diploid chromosome copy number is halved (Haploid).
Meroblastic Cleavage leaves an indentation/cleft because compartmentalizing nascent membranes cannot typically form in isolecithal ova with enriched and dispersed fat (yolk).
Mixotrophs can exploit autotrophic and heterotrophic anabolisms.
mRNA stands for messenger ribonucleic acid which is transcribed in the nucleus from an organism’s active genes before translocation to the cytoplasm where a ribosome translates its sequence and concatenates Amino Acids supplied by transfer ribonucleic acids (tRNAs) into a Polypeptide that later folds into a functional protein. See figure 3. of Pioneering Studies.
Nematosomes are motile multicellular bodies comprising nematocysts (@spirocysts) and flagellated cells of the starlet sea anemone, Nematostella vectensis.
Nuclease enzymes cleave the phosphodiester bonds between the deoxy- or -ribonucleotides of the sugar-phosphate backbone of DNA or RNA but usually only from 5 or 3’ (prime) ends (Russell 1998). The cells of all “living” organisms retain complex DNA proofreading-, damage recognition-, and repair system-competencies. Endonuclease excises a section of ssDNA on either side of damage or aberrations from within a double helix (dsDNA) after which DNA polymerase fills in the gap and ligase seals the sugar-phosphate backbone (Russell 1998a).
Nucleic Acid is typically molecular ribonucleic acid (RNA) or deoxyribonucleic acid (DNA) which maybe complementary (cDNA), double- or single-stranded (dsDNA or ssDNA).
Nucleolus is a dark staining intranuclear body comprising RNA, stored ribosomes, and their auxiliary machinery, which is present during anaphase but degrades during mitosis, yet they are reconstituted following the reorganization of nuclear membranes (Wallace et al. 1996).
Oligonucleotide is a short, typically synthetic Nucleic Acid.
Oligotrophic refers to aquatic environments with few pollutants such as ultralow dissolved inorganic nitrogen (N) and phosphorus (P) akin to pristine reef ecosystems.
Ontogeny/Ontogenesis describes the development of an organism from the moment of fertilization to a mature adult.
Ontology is used contextually to describe the intra- and extra-cellular metabolic pathways associated with an organisms day-to-day housekeeping, homeostatic, and reproductive processes.
Oolemma is an egg’s cellular membrane.
Ooplasm is the cytoplasm of an egg.
Open Reading Frames (ORFs) are the transcriptional parts of genes that lack start and stop sequences but encode for triplet mRNA bases (Codons) which correspond to each kind of Amino Acid.
Orthologue refers to a sequenced and characterized gene that is integral to an elucidated pathway in numerous organisms.
Paralogues are two or more copies derived from a single ancestral gene residing at distal loci in the same genome.
Phenologic refers to the interaction of an organism’s life cycle with its environment (Keith et al. 2016).
Phenotype, see Phenotypic.
Phenotypic Plasticity foregoes the accepted developmental and evolutionary paradigms and expedites swift morphological responses to environmental change.
Phenotypic traits are an organism’s observable characteristics which include how it interacts with its environment that originate from its Epigenotype and Genotype.
Photic Zones are sea surface regions extending to approximately 120 meters that receive sufficient light to support photosynthetic organisms.
Phylogenetic Tree “is a graphic representation of the interrelations and evolutionary history of a group of organisms, indicating the relative order of successive divisions of the line of descent, coincident with speciation events” (Wallace et al. 1996).
Phylogeny facilitates organism classification by comparing the sequences of identical or very similar genes (Homologues).
Planulation refers to the release of free-swimming larvae from brood colonies.
Plasmid is a self-replicating typically circular and mostly prokaryotic extrachromosomal molecule of ss or dsDNA. See: Nucleic Acid.
Spawning – countless marine organisms broadcast their spermatozoa and eggs into the water that later fuse and ultimately transform into planktonic larvae. Corals are gonochoric or hermaphroditic where brood colonies internalize sperm released by others which fertilizes oocytes (eggs) where blastulae and then gastrulae are nurtured until free-swimming planulae are released. Other corals liberate sperm and egg bundles that float leisurely to the surface where they degrade and release their cargo (Wallace et al. 1986), or colonies broadcast eggs and sperm directly whose nuclei fuse into a zygote from which zooplanktonic larvae develop. Several brooding species can raise vegetative larvae after cross-fertilization fails (Aslett 2024).
Spermary or Spermatocyst is a male reproductive organ (gonad) that produces spermatozoa.
Spermatocyte is a spermatogonium-derived cell that undergoes meiosis to generate four haploid tailless spermatids.
Spirocysts, cnidocysts, or nematocysts are the capsular stinging apparatuses of cnidocytes that develop from cnido-, nemato-, or spiro-blasts.
Stomodaeum, foregut.
Thyroid-stimulating hormone (TSH) beta is manufactured by the pituitary gland in response to low serum concentrations of thyroxine and thus regulates the activity of a thyroid gland.
Transcription, see mRNA.
Transcriptome is the sum of all the messenger ribonucleic acid (mRNA) transcribed from an organism’s active genes (@ORFs; Lexico 2019).
Nourishing Vitellarium secreted from vitelline cells enriches yolk which assists and supports embryonic development (Caira 2010).
Zooxanthellate is an organism with symbiotic dinoflagellate “algal”-partners.
Zygotes are formed when the nuclei of a spermatozoan and egg fuse after which additional fertilization cannot occur.
References
Amar, K., O., Chadwick, N., E. & Rinkevich, B. (2007) Coral planulae as dispersion vehicles: Biological properties of larvae released early and late in the season. Mar. Ecol. Prog. Ser. 350, 71-78.
Aslett, C., G. (2024) The Complete Reef Aquarist, for hobbyists, the trade and academics – A Conservation Manual. Aslett, C., G. (ed.). Reef Ranch Publishing Ltd, Leeds, West Yorkshire, UK. https://www.ebay.co.uk/itm/116390701502 p 407.
Attalla, T. (2016) Some Ecological Aspects on coral reefs communities of the Red Sea. PhD thesis, Suez Canal University, Egypt.
Ayre, D., J. & Miller, K., J. (2004) Where do clonal coral larvae go? Adult genotypic diversity conflicts with reproductive effort in the brooding coral Pocillopora damicornis. Marine Ecology Progress Series. 277, 95-105.
Babcock, L. C. (1984) Reproduction and distribution of two species of Goniastrea (Scleractinia) from the Great Barrier Reef Province. Coral Reefs. 2, 187-195.
Babcock, R., C., Bull, G., D., Harrison, P., I., Heyward, A., J., Oliver, J., K., Wallace, C., C. & Willis, B., L. (1986) Synchronous spawnings of 105 scleractinian corals species on the Great Barrier Reef. Marine Biology. 90, 379-394.
Baird, A., H., Guest, J., R. & Willis, B., L. (2009) Systematic and biogeographical patterns in the reproductive biology of scleractinian corals. Annual Review of Ecology, Evolution, and Systematics. 40(1), 551-557. https://www.dio.org/10.1146/annurev.ecolsys.110308.120220
Boch, C., A., Ananthasubramaniam, B., Sweeney, A., M., Doyle, F., J. & Morse, D., E. (2011) Effects of light dynamics on coral spawning synchrony. Biol. Bull. 220, 161-173. https://www.doi.org/10.1086/BBLv220n3p161
Bongaerts P, Cooke IR, Ying H, Wels D, den Haan S, Hernandez-Agreda A, Brunner CA, Dove S, Englebert N, Eyal G, Foret S, Grinblat M, Hay KB, Harii S, Hayward DC, Lin Y, Mihaljevic M, Moya A, Muir P, Sinniger F, Smallhorn-West P, Torda G, Ragan M, A, van Oppen MJ, H & Hoegh-Guldberg O (2021) Morphological stasis masks ecologically divergent coral species on tropical reefs. Curr. Biol. 31(2286-2298), e2288.
Bouwmeester, J., Baird, A., H., Chen, C., J., Guest, J., R., Vicentuan, K., C. & Berumen, M., L. (2014) Multi-species spawning synchrony within scleractinian coral assemblages in the Red Sea. Coral. Reefs. 34, 65-77. https://doi.org/10.1007/S00338-014-1214-6/FIGURES/3
Bouwmeester, J., Berumen, M. L., and Baird, A. H. (2011). Daytime broadcast spawning of Pocillopora verrucosa on coral reefs of the central Red Sea. Galaxea. J. Coral. Reef. Stud. 13, 23–24. doi: 10.3755/galaxea.13.23
Brady, A., K., Hilton, J., D. & Vize, P., D. (2009) Coral spawn timing is a direct response to solar light cycles and is not an entrained circadian response. Coral Reefs 28, 277-280.
Brady, A., K., Snyder, K., A. & Vize, P., D. (2011) Circadian Cycles of Gene Expression in the Coral, Acropora millepora. PLoS ONE 6(9): e25072. https://www.doi.org/10.1371/journal.pone.0025072/
Brady, A., K., Willis, B., L., Harder, L., D. & Vize, P., D. (2016) Lunar Phase Modulates Circadian Gene Expression Cycles in the Broadcast Spawning Coral Acropora millepora. Biol. Bull. 230, 130-142.
Bridge TCL, Cowman PF, Quattrini AM, Bonito VE, Sinniger F, Harii D, Head CEI, Hung JY, Halafihi T, Rongo T & Baird AH (2024) A tenuis relationship: traditional taxonomy obscures systematics and biogeography of the ‘Acropora tenuis’ (Scleractinia: Acroporidae) species complex. Zoological Journal of the Linnean Society. 202, 1-24. https://doi.org/10.1093/zoolinnean/zlad062
Brooke, S. & Järnegren, J. (2013) Reproductive periodicity of the scleractinian coral Lophelia pertusa from the Trondheim Fjord, Norway. Mar. Biol. 160, 139-153.
Bünning, E. & Müller, D. (1961) Wie messen organismen lunare zyklen? Z. Naturforsch. B 16, 391-395. https:///www.doi.org/10.1515/znb-1961-0609
Caira, N., G. (2010) Trypanorhyncha. Planetary Biodiversity Inventory: Survey of Tapeworms from Vertebrate Bowels of the Earth. Illustrated Glossary. https://sites.google.com/site/tapewormpbi/tapeworm-resources/illustrated-glossary
Capel, K., Migotto, A., Zilberberg, C. & Kitahara, M. (2014) Another tool towards invasion? Polyp “bail-out” in Tubastraea coccinea. Coral Reefs. 33(4), 1165-1165.
Crane, K., R. (1999) Reproductive Biology of Scleractinian Coral at Rottnest Island, Western Australia. Murdoch: Honours.
Dale, J., W. (1998) Molcular Genetics of Bacteria Third Edition. Dale, J., W. (ed.). John Wiley & Sons Ltd, Chitchester, West Sussex, UK. pp 286-295.
Davies, T., W., Levy, O., Tidau, S., de Barros Marangoni, L., F., Wiedenmann, J. D’Angelo, C. & Smyth, T. (2023) Global disruption of coral broadcast spawning associated with artificial light at night. Nature Communications. 14, 2511. https://doi.org/10.1038/s41467-023-38070-y
Fadlallah, Y., H. (1983). Sexual reproduction, development and larval biology in scleractinian corals – A review. Coral Reefs. 2, 129-150. https://www.doi.org/10.1007/BF00336720/METRICS
Fan, T. Y., Li, J., J., Le, S., X. & Fang, L., S. (2002) Lunar periodicity of larval release by pocilloporid corals in Southern Taiwan. Zool. Stud. 41, 288-293.
Figueiredo, J., Tomas, C., J., Deleersnijder, E. et al. (2022) Global warming decreases connectivity among coral populations. Nature Climate Change. 12, 83-8.
Fukushiro, M., Takeuchi, T., Takeuchi, Y. et al. (2011) Lunar phase-dependent expression of cryptochrome and a photoperiodic mechanism for lunar phase-recognition in a reef fish, goldlined spinefoot. PLoS One. 6, e28643.
Furukawa M, Kitanobo S, Ohki S, Teramoto MM, Hanahara N & Morita M (2024) Integrative taxonomic analyses reveal that rapid genetic divergence drives Acropora speciation. Molecular Phylogenetics and Evolution. 195, 108063. https://doi.org/10.1016/j.ympev.2024.108063
Gilmour, J., Speed. C., W. & Babcock, T. (2016) Coral reproduction in Western Australia. Peer J. 4, e2010. https://www.dio.org/10.7717/peerj.2010
Gittins, J., R., D’Angelo, C., Oswald, F., Edwards, R., J. & Wiedenmann, J. (2015) Fluorescent protein-mediated colour polymorphism in reef corals: multicopy genes extend the adaptation/acclimatization potential to variable light environments. Mol. Ecol. 24, 453-465.
Guest, J., R., Baird, A., H., Goh, B., P., L. & Chou, L., M. (2002) Multispecific, synchronous coral spawning in Singapore. Coral Reefs. 21(4), 422-423.
Hanafy, M., H., Aamer, M., A., Habib, M., Rouphael, A., B. & Baird, A., H. (2010) Synchronous reproduction of corals in the Red Sea. Coral. Reefs. 29, 119-124. https://www.doi.org/10.1007/S00338-009-0552-2/TABLES/5
Harriott, V., J. (1983) Reproductive ecology of four scleractinian species at Lizard Island, Great Barrier Reef. Coral Reefs. 2(1), 9-18. https://www.dio.org/DOI 10.1007/BF00304727
Harriott, V., J. (1983a) Reproductive seasonality, settlement, and post-settlement mortality of Pocillopora damicornis (Linnaeus), at Lizard Island, Great Barrier Reef. Coral Reefs. 2(3), 151-157. https://www.dio.org/10.1007/BF00336721
Harrison, P., L., Babcock, R., C., Bull, G., D., Oliver, J., K., Wallace, C., C. & Willis, B., L. (1984) Mass spawning in tropical reef corals. Science. 223, 1186-1189.
Hastings, M., H. (1981) The entraining effect of turbulence on the circa-tidal activity rhythm and its semi-lunar modulation in Eurydice pulchra. J. Mar. Biol. Assoc. U. K. 61, 151-160.
Heyward, A., J. & Babcock, R., C. (1986) Self- and cross-fertilization in scleractinian corals. Mar. Biol. 90, 191-195. https://www.doi.org/10.1007/BF00569127
Hunter, C. (1993) Genotypic Variation and Clonal Structure in Coral Populations With Different Disturbance Histories. Evolution. 47(4), 1213-1228.
Huret, J., L., Leonard, C. & Savage, J., R., K. (2000) Chromosomes, Chromosome Anomalies. MRC Radiation, Genome Stability Unit, Harwell, Didcot, OX11 0RD, UK. https://atlasgeneticsoncology.org/teaching/30084/chromosomes-chromosome-anomalies
Jakie, P., L. & York, R., H. (1982) Solar ultraviolet photobiology of the reef coral Pocillopora damicornis and symbiotic zooxanthellae. Bull. mar. Sci. 32, 301-315.
Jokiel, P., L., Ito, R., Y. & Liu, P., M. (1985) Pocillopora damicornis. Marine Biology. 88, 167-174.
Jones, D., A. & Naylor, E. (1970) The swimming rhythm of the sand beach isopod Eurydice Pulchra. J. Exp. Mar. Biol. Ecol. 4, 188-199. https://www.doi.org/10.1016/0022-0981(70)90024-9
Kaiser, T., S. & Neumann, D., J. (2021) Circalunar clocks-old experiments for a new era. Bioessays. 43, e2100074. https:///www.doi.org/10.1002/bies.202100074
Kaniewska, P., Alon, S., Karako-Lampert, S., Hoegh-Guldberg, O. & Levy, O. (2015) Signaling cascades and the importance of moonlight in coral broadcast mass spawning. ELife 4, e09991. https://www.doi.org/10.7554/ELIFE.09991.001
Keith, S., A. et al. (2016) Coral mass spawning predicted by rapid seasonal rise in ocean temperature. Proc. Biol. Sci. 283, 3149-3160.
Kojis, B., L. & Qninn, N., J. (1981) Aspects of sexual reproduction and larval development in the shallow water hermatypic coral Goniastrea australiensis (=favulus) (Edwards and Haime, 1857). Bull. mar. Sci. 3I, 558-573.
Langley C, Harrison PL & Doropoulos C (2024) Pptimizing initial stocking densities of wild coral spawn slicks for mass production of larvae and settled corals for restoration. Restor. Ecol. 32, e14239.
Levitan, D., R., Fukami, H., Jara, J., Kline, D., McGovern, T., M., McGhee, K., E., Swanson, C., A. & Knowlton, N. (2004) Mechanisms of Reproductive Isolation Among Sympatric Broadcast-Spawning Corals of Montastraea annularis species complex. Evolution. 58, 308-323.
Levy, O., Appelbaum, L., Leggat, W., Gothlif, Y., Hayward, D., C., Miller, D. J. et al. (2007) Light-Responsive Cryptochromes from a Simple Multicellular Animal, The Coral Acropora Millepora. Science. 318(5849), 467-70.
Lexico (2019) Powered by Oxford Dictionary. https://www.lexico.com/en/definition/transcriptome
Lin, C., H. & Nozawa, Y. (2023) The influence of seawater temperature on the timing of coral spawning. Coral. Reefs. 42, 417-426. https://www.doi.org/10.1007/S00338-023-02349-9/FIGURES/5
Lin, C-H., Takahashi, S., Mulla, A., J. & Nozawa, Y. (2021) Moonrise timing is key for synchronized spawning in coral Dipsastraea speciosa. PNAS, 118(34), e2101985118.
Linden, B., Huisman, J. & Rinkevich, B. (2018) Circatrigintan instead of lunar periodicity of larval release in a brooding coral species. Nature Scientific Reports. 8, 5668. https://dio.org/10.1038/s41598-018-23274-w
Maxwell, W., G., H. (1968) Atlas of the Great Barrier Reef. Elsevier, New York. p 258.
Mendes, J., M. & Woodley, J., D. (2002) Timing of reproduction in Montastraea annularis: relationship to environmental variables. Mar. Ecol. Prog. Ser. 227, 241-251. https://www.doi.org/10.3354/MEPS227241
Mercier, A., Sun, Z. & Hamel, J-F. (2011) Reproductive periodicity, spawning and development of the deep-sea scleractinian coral Flabellum angulare. Mar. Biol. 158, 371-380.
Merriam Webster (2019) Dictionary. https://www.merriam-webster.com/dictionary/amplicon
Miller, K., J. & Ayre, D., J. (2004) The role of sexual and asexual reproduction in structuring high latitude populations of the reef coral Pocillopora damicornis. Heredity. 92, 557-568.
Million, W., C., Ruggeri, M., O’Donnell, S., Bartels, E., Conn, T., Krediet, C., J. & Kenkel, C., D. (2022) Evidence for adaptive morphological plasticity in the Caribbean coral, Acropora cervicornis. Proceedings of the National Academy of Sciences of the United States of America. 119(49), e2203925119.
Naylor, E. (1996) Crab clockwork: the case for interactive circatidal and circadian oscillators controlling rhythmic locomotor activity of Carcinus maenas. Chronobiol. Int. 13, 153-161. https://www.doi.org/10.3109/07420529609012649
Neumann, D. (2014) “Timing in tidal, semilunar, and lunar rhythms.” Annual, Lunar, and Tidal Clocks: Patterns and Mechanisms of Nature’s Enigmatic Rhythms. H. Numata, H. & Helm, B. (eds.). Springer, Tokyo, Japan. pp 3-24.
Oldach, M., J., Workentine, M., Matz, M., V., Fan, Y-J. & Vize, P., D. (2017) Transcriptome dynamics over a lunar month in a broadcast spawning acroporid coral. Molecular Ecology, 26, 2514-2526.
Olhorst, S. L. (1980) Jamaican coral reefs: important biological and physical parameters. Ph.D. thesis. The University of Yale. p 163.
Oliver, J. & Babcock, R. (1992) Aspects of the fertilization ecology of broadcast spawning corals: sperm dilution effects and in situ measurements of fertilization. Biol. Bull. 183, 409-417. https://www.doi.org/10.2307/1542017
Osman, E., O., Suggett, D., J., Attalla, T., M., Casartelli, M., Cook, N., El-Sadek, I., Gallab, A., Goergen, E., A., Garcias-Bonet, N., Glanz, J., S., Pereira, P., H., Ramirez-Sanchez, M., Santoro, E., P., Stead, A., Yoder, S., Benzoni, F. & Peixoto, R., S. (2024) Spatial variation in spawning timing for multi-species Acropora assemblages in the Red Sea. Front. Mar. Sci. 11, 1333621. https://www.doi.org/10.3389/fmars.2024.1333621
Penland, L., Kloulechad, J., Idip, D. & van Woesik, R. (2004) Coral spawning in the western Pacific Ocean is related to solar insolation: evidence of multiple spawning events in Palau. Coral Reefs. 23(1), 133-140.
Peres, R., Reitzel, A., M., Passamaneck, Y., Afeche, S., C., Cipolla-Neto, J., Marques, A., C. & Martindale, M., Q. (2014) Developmental and light-entrained expression of melatonin and its relationship to the circadian clock in the sea anemone Nematostella vectensis. EvoDevo. 5, 26. http://www.evodevojournal.com/content/5/1/26
Poloczanska, E., Burrows, M., Brown, C., Molinos, J., G., Halpern, B., Hoegh-Guldberg, O. et al. (2016) Responses of marine organisms to climate change across oceans. Front. Mar. Sci. 3, 62. https://www.doi.org/10.3389/fmars.2016.00062
Prasetia, R., Sinniger, F. & Harii. S. (2016) Gametogenesis and fecundity of Acropora tenella (Brook 1892) in a mesophotic coral ecosystem in Okinawa, Japan. Coral Reefs, 35, 53-62. https://www.dio.org/10.1007/s00338-015-1348-1
Pratchett, M., S., Gust, N., Goby, G. & Klanten, S., O. (2001) Consumption of coral propagules represents a significant trophic link between corals and reef fish. Coral Reefs. 20, 13-17. https://www.doi.org/10.1007/s003380000113
Purdy, E. & Winterer, E. (2005) Contradicting Barrier Reef relationships for Darwin’s Evolution of reef types. International Journal of Earth Sciences. 95(1), 143-167.
Ramirez-Portilla C, Baird AH, Cowman PF, Quattrini AM, Harii S, Frederic Sinniger F & Flot J-F (2022) Solving the Coral Species Delimitation Conundrum. Syst. Biol. 71(2), 461-475. http://www.dio.org/10.1093/sysbio/syab077
Revilla-I-Domingo, R., Rajan, V., B., V., Waldherr, M., Prohaczka, G., Musset, H., Orel, L. et al. (2021) Characterization of cephalic and non-cephalic sensory cell types provides insight into joint photo- and mechanoreceptor evolution. eLife 10, e66144. https://www.doi.org/10.7554/eLife.66144
Richmond, R. (1985) Reversible metamorphosis in coral planula larvae. Marine Ecology-Progress Series. 22, 191-185.
Rinkevich, B. & Loya, Y. (1979) The Reproduction of the Red Sea Coral Stylophora pistillata. II. Synchronization in Breeding and Seasonality of Planulae Shedding. Mar. Ecol. Prog. Ser. 1, 145-152.
Rinkevich, B. & Loya, Y. (1979a) The Reproduction of the Red Sea Coral Stylophora pistillata. I. Gonads and Planulae. Mar. Ecol. Prog. Ser. 1, 133-144.
Rinkevich, B. & Loya, Y. (1987) Variability in the pattern of sexual reproduction of the coral Stylophora pistillata at Eilat, Red-Sea: a longterm study. Biol. Bull. 173, 335-344.
Rinsky, M., Aguillon, R., Simon-Blecher, N., Doniger, T., Appelbaum, L. & Levy, O. (2023) CLOCK evolved in cnidaria to synchronize internal rhythms with circadian environmental cues. eLife Evol. Biol. https://doi.org/10.7554/eLife.89499.1
Rock, A., Wilcockson, D. & Last, K., S. (2022) Towards an Understanding of Circatidal Clocks. Front. Physiol. 13, 830107. https://www.doi.org/10.3389/fphys.2022.830107
Ruggles, C., L., N. (1997) Whose Equinox? Journal for the History of Astronomy. 28(22), S44-S50.
Russell, P., J. (1998) Genetics Fifth Edition. Russell, P., J., Buck, E & Dhillon, A. (eds.). The Benjamin/Cumming Publishing Company Inc., Melo Park, CA, USA. p G 8.
Russell, P., J. (1998a) Genetics Fifth Edition. Russell, P., J., Buck, E & Dhillon, A. (eds.). The Benjamin/Cumming Publishing Company Inc., Melo Park, CA, USA. p 637.
Russell, P., J. (1998b) Genetics Fifth Edition. Russell, P., J., Buck, E & Dhillon, A. (eds.). The Benjamin/Cumming Publishing Company Inc., Melo Park, CA, USA. p 694.
Sager, J., C. & Mc Farlane, C, J. (2017) Chapter 1, Radiation. Plant Growth Chamber Handbook. Langhans, R., W. & Tibbitts, T., W. (eds.). Iowa Agriculture and Home Economics Experiment Station Special Report No. 99. North Central Regional Research Publication No. 340. pp 1-30.
Sammarco, P., W. (1982) Polyp Bail-Out: An Escape Response to Environmental Stress and a New Means of Reproduction in Corals. Marine Ecology Progress Series. 10, 57-65.
Schmidt-Roach, S., Miller, K., J., Woolsey, E., Gerlach, G. & Baird, A., H. (2012) Broadcast Spawning by Pocillopora Species on the Great Barrier Reef. PLoS ONE. 7(12), e50847.
Shlesinger, Y. & Loya, Y. (1985) Coral community reproductive patterns: red sea versus the great barrier reef. Science .228, 1333-1335. https://www.doi.org/10.1126/SCIENCE.228.4705.1333
Shoguchi, E., Tanaka, M., Shinzato, C., Kawashima, T. & Satoh, N., A. (2013) Genome-Wide Survey of Photoreceptor and Circadian Genes in the Coral, Acropora Digitifera. Gene. 515(2), 426-431.
Stoddart, J., A. (1984) Genetic differentiation amongst populations of the coral Pocillopora damicornis off southwestern Australia. Coral Reefs. 3(3), 149-156. https://www.dio.org/10.1007/BF00301959
Schuhmacher, H. & Zibrowius, H. (1985) What is hermatypic? Coral Reefs. 4, 1-9.
Szmant-Froelich, A., Riggs, L. & Reutter, M. (1984) Sexual reproduction in Caribbean reef corals. Am. Zool. 23, 961.
Tanner, J., E. (1996) Seasonality and lunar periodicity in the reproduction of Pocilloporid corals. Coral Reefs. 15, 59-66.
Thomson, D. & Frisch, A. (2010) Extraordinarily high coral cover on a nearshore, high-latitude reef in south-west Australia. Coral Reefs 29(4):923-927. https://www.dio.org/10.1007/s00338-010-0650-1
Toledo-Hernandez, C., Ruiz-Diaz, C., P., Ramirez-Lugo, J., S., Torres-Diaz, M., Santiago-Pagan, L., Bruno-Chardon, A. & Diaz-Vazquez, L., M. (2023) Uncovering the link between environmental factors and coral immunity: A study of fluorescent protein expression and phenoloxidase activity in Acropora cervicornis. Forntiers in Marine Scinence. https://dio.org/10.3389/fmars.2023.1133486
van Woesik, R., (2010) Calm before the spawn: global coral spawning patterns are explained by regional wind fields. Proc. R. Soc. B. 277, 715-722. https://www.doi.org/10.1098/rspb.2009.1524
van Woesik, R., Lacharmoise, F. & Köksal, S. (2006) Annual cycles of solar insolation predict spawning times of Caribbean corals. Ecol. Lett. 9, 390-398. https://doi.org/10.1111/J.1461-0248.2006.00886.X
Vize, P. D., Embesi, J., A., Nickell, M., Brown, D., P & Hagman, D., K. (2005) Tight temporal consistency of coral mass spawning at the Flower Garden Banks, Gulf of Mexico, from 1997-2003. Gulf Mex. Sci. 23, 107-114.
Wallace, C., C., Babcock, R., C., Harrison, P., L., Oliver, J., K. & Willis, B., L. (1986) Sex on the reef: mass spawning of corals. Oceanus. 29, 38-42.
Wallace, R., A., Sanders, G., P. & Ferl, R., J. (1996) Biology: The Science of Life, Fourth Edition, HaperCollins Publishers Inc. New York, USA. G-20..
Westneat, M. & Resing, J. (1988) Predation on coral spawn by planktivorous fish. Coral Reefs. 7, 89-92. https://www.doi.org/10.1007/BF00301646
Willis, B., L., Babcock, R., C., Harrison, P., L. & Oliver, J., K. (1985) Patterns in the mass spawning of corals on the Great Barrier Reef from 1981 to 1984. Proc 5th Int Coral Reef Symp. 4, 343-348.
Wilson, M. World Feeds Limited, 3b Coulman Street Industrial Estate, Thorne, Doncaster, DN8 5JS, United Kingdom.
Zakai, D., Dubinsky, Z., Avishai, A., Caaras, T. & Chadwick, N., E. (2006) Lunar periodicity of planula release in the reef-building coral Stylophora pistillata. Mar. Ecol. Prog. Ser. 311, 93-102.
Zhang, L., Hastings, M., H., Green, E., W., Tauber, E., Sladek, M., Webster, S., G. et al. (2013) Dissociation of circadian and circatidal timekeeping in the marine crustacean Eurydice pulchra. Curr. Biol. 23, 1863-1873. https://www.doi.org/10.1016/j.cub.2013.08.038
For more information, visit https://reefranch.co.uk/
![]()

0 Comments