



Corals of the orders Rugosa and Tabulata were replaced by ancient reef-forming (“hard rayed”) Scleractinia around two hundred and fifty million years ago as animals began to inhabit the land, which originated from an earlier distantly-related anemone-like ancestor (Fig 24.; Waggoner et al. 1994) yet their evolution to present-day hermatypes coincided with the rise of their dinoflagellate symbionts (zooxanthellae) ninety million years later. The Permian/Triassic mass extinction had delayed their evolution forty million years before insofar as it terminated the epoch’s primordial photosynthetic endosymbiosis (LaJeunesse et al. 2018.
Recent molecular analyses discovered that Scleractinia are descended from two distinct ancestors that diverged over two hundred million years ago (Romano & Palumbi 1996; Fukami et al. 2008; Budd et al. 2010; Kitahara et al. 2010; Kayal et al. 2013; Okubo et al. 2013). These Robusta and Complexa lineages comprise corals that construct high- and low-density massive boulder-like/plating and branching edifices akin to the genera Goniastrea and Acropora (above). Of note is the Robusta family Pocilloporidae (Kitahara et al. 2010) that comprises branching Pocillopora damicornis. Comprehensive DNA and mitochondrial 16 svedberg (16S) ribosomal ribonucleic acid (rRNA) sequence analyses (Romano & Palumbi 1996) discovered that the hitherto considered monophyletic ancestries of reef-forming corals where skeletal morphology and the fossil record heavily influenced species discernment, were polyphyletic which demanded the revaluation of canonical taxonomic ranks and phylogenetic trees (Kitahara et al. 2010; Ying et al. 2018). Therefore, much of hard coral classification remains uncertain (incertae sedis) and is undergoing extensive revision (Huang et al. 2011; Okubo et al. 2013). Nevertheless, the findings of Kitahara and colleagues in 2010 substantiated the monophyly of the families Acroporidae, Poritidae, Pocilloporidae, and Fungiidae if the genus Alveopora is moved from Poritidae to Acroporidae. The remaining poritid genera: Goniopora and Porites would form a monophyletic clade while the acroporid genus Anacropora would continue as a sister taxon of Acropora, Isopora, and Montipora (Wallace et al. 2007; Kitahara et al. 2010).
Unfamiliar terms are clarified in a Glossary in Appendix I, while the author foresees such synonym-derived ambiguity will lead to confusion in retail outlets, publications, or on web forums. Coral genus often impacts retail pricing which is revised throughout taxonomic reshuffling while classification remains in flux.
-

- Fig 1. 12-tentacled polyps of the genus Acropora and family Acroporidae (left; Scleractinia: Hexacorallia)
-

- and an eight-tentacled clove polyp of the “soft” coral genus Clavularia and family Clavulariidae (Malacalcyonacea: Octocorallia).
Why is taxonomy important?
Comprehension of water chemistry and system dynamics is crucial for mariculture where resources like Aslett 2024 provide laypersons with vital insight (https://www.ebay.co.uk/itm/116390701502). However, common names are ambiguous which generally refer to genera, and scientists must perform analyses on the correct organism. Aetiological agents (pathogens) are classified using the same tenets therefore taxonomy facilitates the identification of efficacious remedies, and organisms sharing ancestral lineage are likely to require analogous husbandry and may be sources of compounds with medicinal properties. Taxonomy expedites and supports scientific inquiry and expands our skillset by promoting a greater understanding of coral ecology and the conditions within which they evolved.
There are many successful reef aquarists with breathtaking displays who competently care for most hermatypes without conceiving their names or their evolutionary trajectories, yet equally, there are reef enthusiasts who effortlessly identify species but find their captive culture challenging. And naturally, everyone in between.
Naming and categorizing plants and fungi became necessary early on in human cultures insofar as folk taxonomy categorized what was edible, inedible, poisonous, and healing. The Chinese and Egyptians documented species from 3,000 BC and 1,500 BC, whereas various forms of life were recorded by the Greeks and Romans from 400 BC to 80 AD (Manktelow 2010). Such information remained unavailable and could not influence scientific investigations until the Middle Ages.
-

- Fig 2. A mushroom of the genus Ricordea and family Ricordeidae (left; Hexacorallia: Corallimorpharia)
-

- and “soft” polyps of the genus Palythoa and family Sphenopidae (Hexacorallia: Zoantharia).

Fig 3. Black coral, Antipathes cf. grandis where the findings of Kitahara and colleagues suggested in 2010 that the order Antipatheria did not radiate from within the subclass Hexacorallia.
The 16th century’s optical lenses assisted the scrutiny of magnified botanical samples by pioneers like the Bauhin brothers, Caesalpino, John Ray, and Joseph Pitton de Tournefort. Carl Linnaeus deemed the father of modern taxonomy classified plant and zoological specimens in the 18th century, whereas the French taxonomists Antoine Laurent de Jussieu, Baptiste de Lamarck, and Georges-Luise Leclerc de Buffon recognised heritable traits and introduced the concept of evolution and established class, family, and genus which heavily influenced Charles Darwin. However, the inevitable inconsistencies of international nomenclature necessitated the development of a global framework (Manktelow 2010).
-

- Fig 4. Toadstool coral, Sarcophyton species (left; Octocorallia: Alcyonacea)
-

- and star polyps, Briareum species (Octocorallia: Alcyonacea).
Darwin’s theory of natural selection motivated efforts to rank species according to ancestral lineage when the first evolutionary trees were constructed in Germany by Ernst Haeckel and August Wilhelm Eichle in the 19th and 20th centuries. Haeckel devised a process of determining divergent evolution arising from a mutual ancestor termed phylogeny, and elucidation of the genetic code by Watson and Crick in the 1950s sanctioned classification according to chromosomes, biochemistry, molecular biology, and proteins (Watson & Crick 1953; Pray 2008; Manktelow 2010). DNA sequence analyses of the 1980s facilitated discernment by comparing similar genes (homologues), whilst any system had to accommodate clades comprising all the descendants from a common ancestor (monophyly; Manktelow 2010).

Fig 5. A sea fan of the subclass Octocorallia previously ranked in the orders Alcyonacea and Gorgonacea.
Species are assigned kingdom, phylum, class, order, family, and genus, and subdivisions and clades occur frequently. Genus and species are italicised while the first letter of the former and higher classifications are UPPERCASE, whereas the entire ancestries of viruses are italicised. Subspecies like Photobacterium damsela damsela and a year and classifying scientist(s) add a further layer of specificity like Acropora millepora Ehrenberg 1834 (WoRMS 2020).

Fig 6. A brain coral, Colpophyllia natans (Scleractinia: Hexacorallia) integral to the Floridian coral rescue initiative. Image courtesy of The Florida Fish and Wildlife Commission ©.

Fig 7. A diagram of a eukaryotic animal cell featuring its organelles.
The taxological search engine, The World Register of Marine Species (WoRMS; http://www.marinespecies.org/) is favored by the author which is less cluttered and clearer than The National Centre for Biotechnology Information’s (NCBI’s) taxonomy browser “root” (https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/). Furthermore, the former tends to recognize and accept contemporary findings in comparison to the latter. For example, much of the species-rich phagocytic mixotrophic chromists of which the infrakingdom Heterokonta is conspicuous, were hitherto considered Protozoa of the kingdom Protista (Cavalier-Smith et al. 2006; Cavalier-Smith 2010; Cavalier-Smith 2018), yet the NCBI is yet to accept and adopt Cavalier-Smith and colleagues’ recommendations. Many researchers do not recognize WoRMS classifications because of the NCBI’s elite criteria, while table 1. features the taxonomically accurate genus or species of 65 popular captive corals accepted by WoRMS in 2022 (Aslett 2024). The author’s aim was to present taxonomically definitive data, albeit taxonomy continues to evolve, the validity of each historic rank is associated with a year.

Fig 8. Polypeptide fabrication: transcription (expression) of intranuclear deoxyribonucleic acid (DNA) creates messenger ribonucleic acid (mRNA) which is translated by a ribosome (pink) after translocation to the cytoplasm. Amino acids bound to transfer ribonucleic acids (tRNAs) are concatenated according to the mRNA sequence, while their R-group (sidechain) charges and shapes determine protein folds.

Fig 9. A phylogenetic tree of the descent of anthozoan clades where each subsequent branch represents divergent evolution. Adapted from Kitahara et al. 2010.

Fig 10. A phylogenetic tree of the families of the scleractinian Basal and Complexa clades adapted from Kitahara et al. 2010 (Fig 9.).
Corals are diploid insofar as their nuclei comprise pairs of chromosomes (2n) each originating from one parent that encode for typically distinct forms of genes called alleles. Each spermatozoan or egg (ovum; oocyte) is haploid (1n) whose mitochondria remain maternally inherited because they are equipped with their own circular DNA genome akin to chloroplasts (@ plastids; Russell 1998a). Intracellular organelles likely arose from primordial endosymbiosis (Janouškovec et al. 2010; Bonacolta et al. 2023) while genes encoding ribosomal proteins have been identified in mitochondria and plastids (Maier et al. 2013). Therefore, a single ancestral cell may have accommodated up to four kinds of protein-synthesizing ribosomes. Phylogenetic sequence analyses of these laterally transferred rRNA-encoding genes may thus elucidate the origins of Eukaryota (Figs 7. & 8.; Martin et al. 2015).
The endosymbiotic paradigm was conceived in the 19th century. Microscopy and the eminent deductions of the day proposed developmental reduction of a cyanobacterium led to the ascent of plastids inhabiting a heterotrophic host which descended from an amoeba that engulfed and cohabited with a micrococcoid mutualist responsible for the nucleus of eukaryotes (Mereschkowsky 1905, cited in Martin et al. 2015). The origins of the eukaryotic nucleus, intracellular cytoskeleton, flagella, and mitochondria remain enigmatic with theories too numerous to include (Martin et al. 2015). Nevertheless, Allen’s co-location for redox regulation (CoRR) hypothesis rationalizes why mitochondria and chloroplasts have retained genomes and the ribosomal proteins that express them, because they sustain their electron transport chains (Allen 1993; Allen 2003). Furthermore, the genes encoding their ribosomes have evolved convergently while their intra-compartmental co-expression is a prerequisite for their biogenesis (Maier et al. 2013). Hence studies of mitochondrial rRNA or rDNA assist elucidation of maternal ancestry.

Fig 11. [Bi] a phylogenetic tree of the families of the significantly polyphyletic scleractinian Robusta clade reported by Kitahara et al. 2010.
WoRMS ranks the coral Acropora millepora in the kingdom Animalia, the phylum Cnidaria, the class Anthozoa, the subclass Hexacorallia, the order Scleractinia, the family Acroporidae, the genus Acropora, and the species millepora (Worms 2020). Corals and humans (Homo sapiens) belong to the same kingdom, yet from there all similarity ends because the former have been around much longer and rapidly adapted to their environment.
Animals belonging to the phylum Cnidaria such as corals, anemones, jellyfish, and hydroids have specialized cells called cnidocytes that contain venomous nematocysts (Technau & Steele 2011; Beckmann & Özbek 2012).
The class Anthozoa includes the five hundred and fifty million years old polyp morphotypes: stony corals, anemones, sea fans, and sea pens, with a tube-like body terminated by a disc of stinging tentacles surrounding a mouth. “Soft” corals exploit supportive proteins and mesoglea whereas hard corals have calciferous skeletons (Daly et al. 2006).
The informal classification referring to corals that are large polyp stony (LPS) and small polyp stony (SPS) appeared to originate in the hobby where the former has polyps that may be centimeters across, while those of SPS corals are approximately 2 millimeters in diameter.
Recent sequence analyses of mitochondrial genomes affirmed the class Anthozoa includes the subclass Hexacorallia consisting of members with multiple sums of six tentacles that arise from the orders Actiniaria like starlet anemones (Nematostella vectensis), Antipatharia like black corals (Antipathes grandis), Corallimorpharia like mushrooms (Ricordea florida, Discosoma, and Rhodactis species; spp.), stony Scleractinia like Acropora spp., Colpophyllia spp., Montipora spp., and Pocillopora spp., and Zoantharia such as “soft” polyps (Palythoa spp., Parazoanthus spp., and Zoanthus spp.). The “soft” anthozoan subclass Octocorallia comprises eight tentacled members of the order Alcyonacea like carnation coral (Dendronephthya spp.), toadstool coral (Sarcophyton spp.), star polyps (Briareum spp.), sea whips (Gorgonia spp.), and sea pens, whereas the orders Helioporacea or Pennatulacea include Heliopora coerulea or Stylatula elongata (Figs 1. to 6. & 13.; Daly et al. 2006; Fabricius 2011; Kayal et al. 2013; Malakhov 2016).
Chalice corals are an unofficial scleractinian subgroup of the remarkably aggressive suborder Faviina (subjective synonym: Pectiniidae) which are typically described as cup corals, while their genera include Echinomorpha (Lobophylliidae), Echinophyllia (Lobophylliidae), Echinopora (Merulinidae), Mycedium (Merulinidae), Oxypora (Lobophylliidae), Paraechinophyllia (Lobophylliidae), Pectinia (Merulinidae), and Physophyllia (Merulinidae; Table 1.; Adams 2018; Morice 2022).
Likewise, the casual and marginally paraphyletic clade, Bigmessidae comprises the families Faviidae, Merulinidae, Pectiniidae, and Trachyphylliidae which were somewhat resolved by Huang and allies in 2011

Fig 12. [Bii] a phylogenetic tree of the remaining families of the scleractinian Robusta clade reported by Kitahara et al. 2010.
Every phylogenetic study and tree aim to define the order and line of descent congruent with the evolutionary relationships of a group of organisms (Wallace et al. 1996). Such precision necessitates the identification of monophyletic clades constituting all the lineages radiating from a shared ancestor.
Optical and electronic microscopy facilitated the visual appraisal of biological specimens and fossils upon which taxonomy was founded. However, it became possible to trace the evolution of homologous genes following the dawn of DNA sequence analyses which intuitively arms us with an unequivocal means to discern species ancestries.
Nevertheless, science continues to reveal assumptions muddy the waters of objective studies. Furthermore, attempting to define the evolutionary relationships of optimized eukaryotes by analyzing a handful of homologues whose homeostasis exploits thousands-upon-thousands of genes, must continue to yield inconsistencies.

Fig 13. Plankton-demanding ahermatypic yellow polyps of the genus Parazoanthus and family Parazoanthidae (Hexacorallia: Zoantheria) are one of the few ornamental “soft” corals consumed by peppermint shrimp (Lysmata wurdemanni).

Fig 14. A photograph of an ancient massive yellow scroll coral (Turbinaria reniformis) of the family Dendrophylliidae (Basal: Scleractinia) captured in Egypt’s Red Sea.
To this distinguishing end, coral taxonomists have strived to unearth distinct and conserved intracellular pathways that define each taxon because databases and few sequenced and annotated anthozoan genomes constrain the scope of molecular techniques. Even so, recent advancements in data analyses and the expansion of bioinformatics signal the verge of a significant breakthrough. However, molecular techniques have yielded ordinal and familiar inconsistencies (Romano & Palumbi 1996; Romano & Cairns 2000, cited in Kitahara et al. 2010).
Mitochondrial genotyping (mitotyping) has been the focus of much research insofar as their simple short nucleic acids are economical to sequence while their genes appear highly conserved. Mitochondrial cytochrome oxidase is an indispensable respiratory enzyme whose subunit 1 (CO1) was partially sequenced and used to trace the ancestries of 65 and 11 deep- and shallow-water Scleractinia which identified significant polyphyly in the 25 families of extant euphotic-dwelling reef-forming species. Excluding Caryophylliidae, monophyly pervaded the families of deep-water azooxanthellates such as Dendrophylliidae, Flabellidae, Fungiacyathidae, Gardineriidae, Micrabaciidae, and Turbinoliidae, whereas most zooxanthellates were polyphyletic apart from Acroporidae, Fungiidae, Pocilloporidae, and Poritidae. Greater environmental stability at significant depth may obviate the necessity for the rapid adaptation competency of phenotypic plasticity that confounds expression analyses (Figs 9. to 12.; Kitahara et al. 2010). Nonetheless, DNA sequencing would overcome these disparities, albeit prone to the noise produced by dormant or vestigial homologues, the concise genomes of organelles are parsimonious. Moreover, predominantly non-colonial (solitary) deep-water azooxanthellates of Basal Scleractinia diverged before the radiations of Complexa and Robusta (Fig 9.; Kitahara et al. 2010).

Fig 15. A phylogenetic tree illustrating the revised classifications of the phylum Cnidaria from 2013 discerned by comparing mitochondrial proteins. Branch-associated support values correspond to the probabilities of CATGTR Bayesian analyses/GTR Bayesian analyses/GTR maximum likelihood bootstraps, while asterisks and hyphens represent maximum confidence and marginal inconsistencies. For scale bar see Appendix I. Analyses and evolutionary tree courtesy of Kayal et al. 2013 and the Creative Commons Attribution Licence 2.0.

Table 1. Featuring the taxologically accurate nomenclature of 65 commonly imported corals accepted by WoRMS in 2022 (Aslett 2024).
Mitochondrial CO1 genes do not contain insertions or deletions (indels) like their 12, 16, and 28S rDNA. Remarkably, Bayesian analyses refuted the naked coral hypothesis (Medina et al. 2006) that proposed Corallimorpharia evolved from Scleractinia by means of skeletal loss throughout periods of acute environmental disruption (Fukami et al. 2008; Budd et al. 2010; Kitahara et al. 2010), while the divergence of Robusta occurs from within Agariciidae of Complexa which contrasts to previous studies where ribosomal nucleic acid implied a profound split (Fig 16.; Veron et al. 1986, cited in Kitahara et al. 2010; Chen et al. 1995; Cuif et al. 2003; Fukami et al. 2008; Ying et al. 2018). All Robusta families were significantly para- or poly-phyletic excluding Pocilloporidae. The families Astrocoeniidae; polyphyletic (pp) Caryophylliidae; Euphylliidae; Meandrinidae; pp Oculinidae, and Siderastreidae are distributed across Complexa and Robusta, while the former or latter include: Acroporidae; pp Agariciidae; Dendrophylliidae; Flabellidae; Fungiacyathidae; pp Poritidae; Turbinoliidae or pp Anthemiphyllidae; pp Faviidae; Fungiidae; pp Merulinidae; pp Mussidae; pp Pectiniidae; Rhizangiidae; Stenocyathidae, and Trachyphylliidae (Figs 9. to 12.; Kitahara et al. 2010).
Deepsea solitary azooxanthellate Gardineriidae and Micrabaciidae are the most basal scleractinian groups which are likely analogues of the ancestors of present-day euphotic-dwelling hermatypes (Stolarski & Vertino 2007; Kitahara et al. 2010). Hence these early Scleractinia may have evolved alongside corals of the order Rugosa which perished during the Permian/Triassic mass extinction (Kitahara et al. 2010). Stanley Junior and Swart suggested endosymbiosis arose in the Triassic (Stanley Jr. & Swart 1995) when evidence suggests that primitive Scleractinia inhabited shallow water (Stolarski & Vertino 2007; Kitahara et al. 2010).
Dendrophylliidae and Poritidae share ancestry where the fossil record suggests they emerged from the early- and mid-Triassic respectively (Wells 1956, cited in Kitahara et al. 2010) where Actinacididae may be the direct progenitor of either poritids (Veron 1995, cited in Kitahara et al. 2010) or less likely dendrophylliids (Cairns 2001).
Apart from Pachyseris speciosa which would be better positioned in Euphyllidae, the genera Agaricia, Gardineroseris, and Pavona constitute monophyletic Agariciidae (Kitahara et al. 2010).
If Stylocoeniella guentheri (Robusta: Astrocoeniidae) is nestled within Pocilloporidae, it forms a convincing clade with its congeners Pocillopora, Stylophora, Seriatopora, and Madracis that diverged from the base of the Robusta branch (Kitahara et al. 2010).
Research supports the reclassification of the oculinid Complexa and Robusta genera Ctenella and Galaxea to Euphyllidae, while the extensively diverse families Caryophylliidae; Euphylliidae; Faviidae; Mussidae, and Oculinidae may form a novel clade. Most taxonomic studies appear to infer that geographically and bathymetrically separated species have evolved independently which challenges clades that integrate corals from various depths and worldwide locations (Fukami et al. 2008; Kitahara et al. 2010).
Gene sequence analyses of mitochondrial CO1 and cytochrome oxidase b (cytb), and nucleic b-tubulin and 28S rDNA from 17 scleractinian families, 75 genera, and 127 species from The Pacific and Atlantic, affirmed that 16 families are polyphyletic, where meticulous morphological and molecular studies showed merit for definitive species discernment (Fukami et al. 2008).

Fig 16. [a] a phylogenetic tree nesting Goniastrea and Fungia spp. in Robusta (green) in contrast to complex Acropora, Galaxea, and Porites spp. (blue). A colony (1), the large polyp(s) (2), and corallite(s) (3) of [B] Goniastrea aspera (Robusta); [C] Fungia fungites (Robusta), and [D] Galaxea fascicularis (Complexa). [E] Acropora millepora (Complexa); [F] Acropora digitifera (Complexa); [G] Porites lutea (Complexa), and the anemones: [H] Exaiptasia spp. (synonym: Aiptasia) and [I] Nematostella vectensis. For scale bar see Appendix I. Tree and images courtesy of Ying et al. 2018 and the Creative Commons Attribution Licence 4.0.

Fig 17. Embryonic development from prawn-chip/cushion blastulae to teardrop/elongate gastrulae in “complex” and “robust” scleractinian clades. The former’s blastulae are stereo and solid lacking a blastocoel void. Cell bilayers shrink, thicken, and roll inward forming a bowl-like stage whereafter the cells in the concavity are overgrown by ectodermis. The undifferentiated cells and yolk granules of the core are remodelled into endodermis enveloped in a mesogleal sphere [purple]. The blastopore remains closed through spheroidal gastrulation whereafter the mouth is formed through an invagination of the animal pole (Okubo & Motokawa 2008). “Robust” blastulae are hollow with a pronounced blastocoel void into which no material is excreted while “pseudo-blastopores” [+] close during spheroidal gastrulation whereafter a conventional blastopore materialises, remains open, and eventually forms the mouth during which the blastocoel is filled by cellular ingress. Drawings courtesy of Okubo et al. 2013 and the Creative Commons Attribution License.
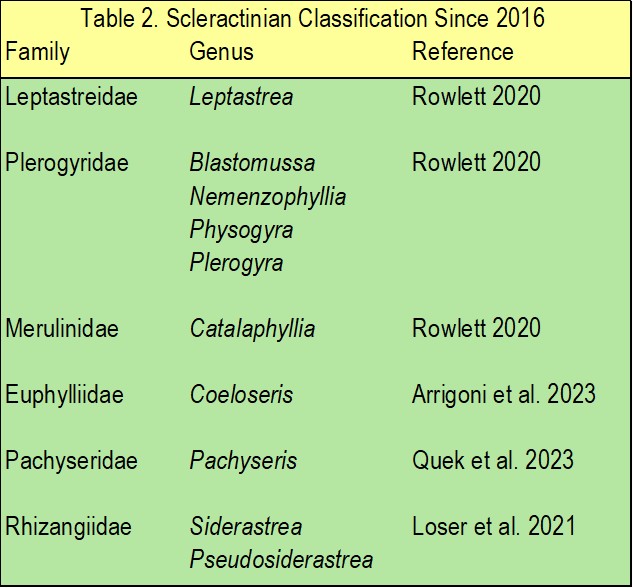
Table 2. Revisions of scleractinian families and genera since 2016 as reported by Fukami and Kitano in 2023.
Fukami and collaborators found the families Faviidae and Trachyphylliidae invalid and that Fungiacyathidae is not a congener of Corallimorpharia in 2008. Mussidae must therefore accommodate most Atlantic faviids excluding polyphyletic Montastraea. They recommended that the faviid genera Cladocora and Solenastrea be absorbed by Oculinidae and species of Leptastrea by Fungiidae which appeared to attain monophyly (Fukami et al. 2008). Kitahara and allies ranked Cladocora in Caryophylliidae while Solenastrea remained in Faviidae whereas the fungiid genus Oulastrea was moved to Faviidae in 2010. Conversely, the family’s significantly polyphyletic Robusta genera: Diploastrea and Plesiastrea were upheld (Kitahara et al. 2010). The questionable species of Mussidae: Acanthastrea and Blastomussa required re-evaluation in 2008 (Fukami et al. 2008), yet their rank was maintained in 2010 (Kitahara et al. 2010).
Polyphyletic Siderastreidae had three monophyletic genera in 2008: Acropora, Porites, and Siderastrea with representatives from The Atlantic and Pacific, whereas Pacific Coscinaraea and Psammocora would be better positioned in Fungiidae (Fukami et al. 2008), but their ancestry remained unchanged in 2010 (Kitahara et al. 2010).
Astrocoeniidae comprises two divergent clades: one adopting Stephanocoenia with no sister taxa, and the other comprising Madracis and Stylocoeniella which share descent with Pocilloporidae. The family was founded by Koby in 1890 using the fossil morphology of Astrocoenia akin to that of a Stephanocoenia (Fukami et al. 2008). Merely Madracis had migrated to monophyletic Pocilloporidae by 2010 (Kitahara et al. 2010).
Euphylliidae acquired Galaxea from Oculinidae and Ctenella from Meandrinidae in 2008 (Fukami et al. 2008), whereas none of these revisions were accepted by Kitahara and collaborators in 2010. Oculinidae remains a sister taxon of Agariciidae with at least one species of Pachyseris. Fukami and colleagues recommended that Alveopora be absorbed into Acroporidae from Poritidae in 2008 which remained unsanctioned by Kitahara and allies in 2010, while the former team’s findings found both families to be non-monophyletic. Dendrophylliidae was one of the few families whose affiliates remained stable while the findings of other studies contested the affinity of Scleractinia with Corallimorpharia (Fukami et al. 2008), like the mitochondrial protein analyses of Kayal and collaborators from 2013 (Fig 15.). Although several stony coral familiar ranks remain uncertain, mitochondrial and nuclear gene pair analyses suggested that the order Scleractinia is a monophyletic clade descended from a common ancestor (Fukami et al. 2008).
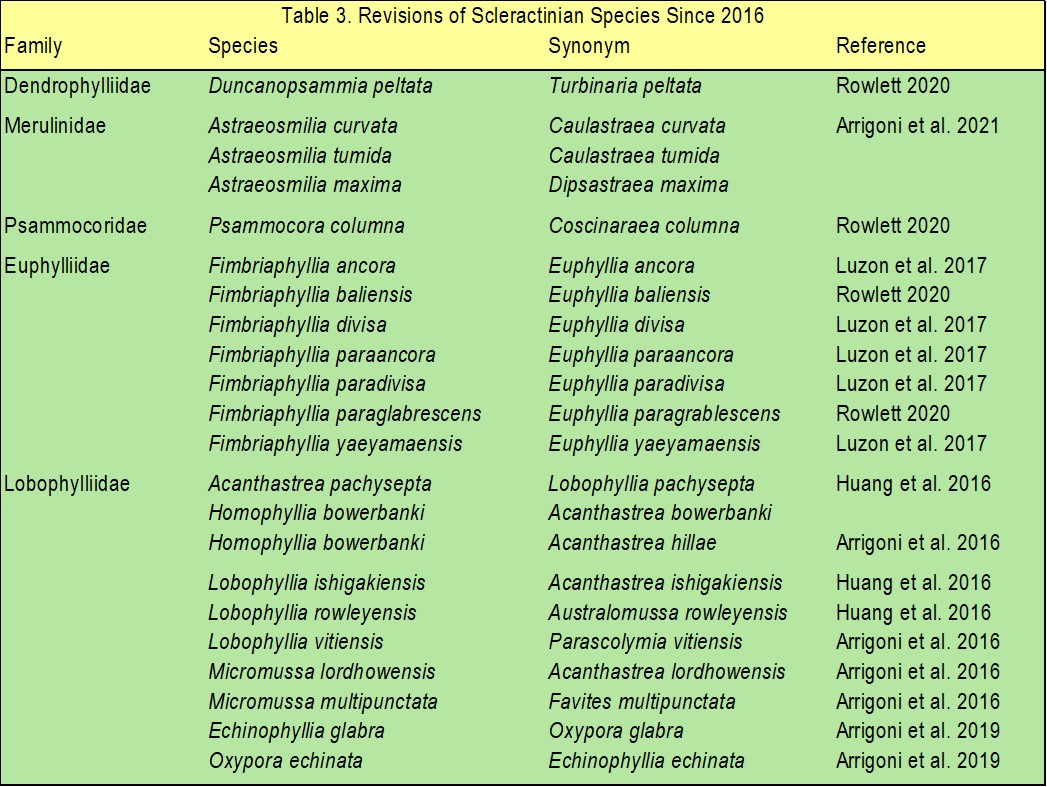
Table 3. Revisions of scleractinian families and species since 2016 as reported by Fukami and Kitano in 2023.
The Robusta subclade, XVII informally named Bigmessidae comprised 132 species mostly from Indo-Pacific Faviidae, Merulinidae, Pectiniidae, and Trachyphylliidae. Merely Trachyphyllia geoffroyi was ranked in the latter (Kitahara et al. 2010) whereas Merulinidae were pervaded with polyphyly. However, equally divergent Faviidae and Pectiniidae included species from within and without XVII (Huang et al. 2011). Members from other families such as Mussidae were also distributed amongst Bigmessidae while genera like Echinopora, Favia, Favites, Goniastrea, Montastraea (Huang et al. 2009), Leptoria, Platygyra, and Oulophyllia were polyphyletic (Fukami et al. 2008).
Huang and collaborators thus ventured to resolve the ancestries of 76 and seven species from within and without Bigmessidae with combined genetic analyses (Huang et al. 2011).
Corals were sampled from the Caribbean, the Philippines, The Great Barrier Reef (GBR), Singapore, and Japan where the laboratory observation of numerous skeletal traits rigorously verified each species. The loci of three nuclear and two mitochondrial sequence biomarkers from each coral were amplified using the polymerase chain reaction (PCR). Nuclear 28S D1 and D2 fragments of rDNA, histone H3, and 5.8S rDNA-integral internally transcribed spacers 1 and 2 (ITS) contrasted with mitochondrial CO1 and their noncoding intergenic region situated between COI and their formylmethionine transfer RNA (tRNA) genes (IGR). It was predicted that classifications arising from these combined sequences would prove definitive (Huang et al. 2011).
Remarkably, nuclear DNA proved more taxonomically-discerning than mitochondrial, where sequences encoding chromosomal DNA-organising and -coiling histone H3 proteins conferred significant likelihood for most Bigmessidae subclades apart from XVII D/E (Fig 18.; Huang et al. 2011). IGRs were so significantly divergent amongst the 83 species they only aligned and proved discriminating for a fraction of clade XVII. Coral genomes encode for several ITS rDNAs but the PCR primers were designed to anneal to merely one copy of each, insofar as amplification of multiple kinds would confound objectivity. Two ITS regions were thus sequenced for each coral that could not be classified using the other four loci; however, the experimenters could not rule out cryptic speciation where distinct species of coral remain outwardly alike. It was postulated that the confidence for nodes determined from the secondary structure analyses of ITS2 encoded on 5.8S rDNA would be unequivocal compared to conventional ITS alignment which was not the case. The ITS sequences of Hydnophora microconos, H. pilosa, and Merulina scabricula would not amplify and were thus excluded. Moreover, not all CO1 regions contributed because they barely differed between species (Huang et al. 2011).
This study was the first to discriminate and rank the species featured in table 4. while the combined and clade-specific phylogenetic trees are represented in figures 18. and 19. (Huang et al. 2011). The comprehensive tree comprises the well-defined Bigmessidae subclades XVII A to I as described by Budd and Stolarski in 2011, and the inferred corroborative outgroups clades XIV to XVI and XVIII to XXI. However, the confidence for the nodes of XVII’s subclades A, C, F and I remained low due to merely <50 of percent of 1,000 bootstraps (Huang et al. 2011).
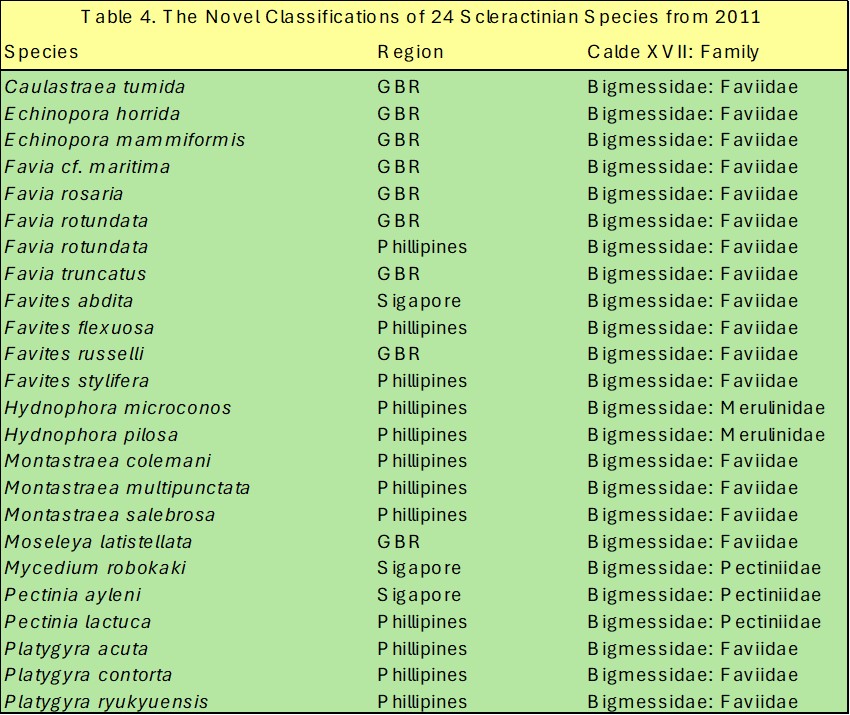
Table 4. The novel clade XVII and familiar rankings of 24 coral species first defined using the combined sequence analyses of three nuclear and two mitochondrial DNA loci reported by Huang and allies in 2011.
Diploastrea heliopora of clade XV and Montastraea cavernosa of XVI sequentially diverge in Fukami and allies’ tree from 2008 while Huang and colleagues suggested they form a mutual split (Fig 18.).
The combined monophyly of clades XVII, XIX, and XX for Pacific Faviidae and Mussidae was weakly upheld by Huang and collaborators in 2011. Montastraea multipunctata and Moseleya latistellata were clade XVII faviids by virtue of their Pacific origins although morphological studies had hitherto ranked them with Acanthastrea hillae and Blastomussa merleti from respective clades XIV (Hodgson 1985) and XVIII (Veron et al. 1977; Veron 1986, cited in Huang et al. 2011). However, M. multipunctata and M. latistellata were classified in clades XIX and XX in 2011. The unambiguous rank of the faviid Diploastrea heliopora of clade XVII is perhaps the only monophyletic member which recommends that most other Pacific faviids be re- or -evaluated including the currently unsampled genera Australogyra, Boninastrea, Erythrastrea, and Paraclavarina (Huang et al. 2011).
The pectiniid genera Pectinia and Mycedium of XVII E are sister taxa of Oulophyllia spp. of XVII D hence the XVII D/E grouping (Huang et al. 2011). The latter genus is monophyletic while species of Caulastraea appear to have descended from a prior divergence and not as Fukami and colleagues suggested in 2008, from a mutual split within XVII A (Huang et al. 2011).
Merulinid Hydnophora are more allied with Favites and Pacific Montastraea than their familiar members Merulina and Scapophyllia. Significantly para- or poly-phyletic Goniastrea, Merulina, and Scapophyllia appear members of XVII A yet the former also appears in B (Huang et al. 2011).
Trachyphylliidae like Trachyphyllia geoffroyi are nested in clade XVII B which is dominated by significantly para- and poly-phyletic species of Favia where Favia lizardensis and F. truncatus are Trachyphyllia geoffroyi’s sister taxa because they radiate from a single branch (Figs 18. & 19.; Huang et al. 2011).
Clade XVII is undergoing extensive taxonomic and nomenclative revision for several reasons least of all, because the faviid Favia fragum is a clade XXI affiliate (Fukami et al. 2008; Huang et al. 2011).
Montastraea annularis, M. faveolata, and M. franksi are the only Atlantic species in clade XVII while the former and the genus Cyphastrea form the well-supported clade XVII C. Nevertheless, they cannot be reconciled using skeletal morphological and deposition traits (Huang et al. 2011), yet procreative strategies and sexual mode are highly conserved which could be used in combination with molecular analyses to boost resolution (Baird et al. 2009; Kerr et al. 2011). Nevertheless, skeletal morphology and optical and electronic microscopy remain phylogenetically-discriminating key procedures, but they do not account for phenotypic plasticity or cryptic speciation (Budd & Stolarski 2009).
Huang a allies demonstrated in 2011 that regionally-proximal yet geographically separated conspecific corals from the Indo-Pacific evolved from several distinct lineages. Australia’s Goniastrea australensis is clustered with Favites russelli and Montastraea curta in figure 18.; however, the Singaporean strains S098 and S099 ranked in subclade XVII A of figure 19., most likely represent hitherto unclassified isolates. Great Barrier Reef Montastraea valenciennesi (G61904) and its counterparts from Singapore (S006 and S008) are ranked in subclade XVII B that is inundated with species of Favia, yet P102 from the Philippines is nested within the youthful subclade XVII F alongside species of Favites (Huang et al. 2011), whereas two genotypically-distinct M. valenciennesi coexist in Japanese waters off Wakayama (Fig 19.; Fukami & Nomura 2009). Hence the Australian and Singaporean isolates could be reclassified as Favia valenciennesi while P102 may be a novel species (Huang et al. 2011). Nevertheless, Favia speciosa and Montastrea valenciennesi had been reclassified as Dipsastraea speciosa and Phymastrea valenciennesi by 2013 (Okubo et al. 2013).

Fig 18. The combined maximum likelihood phylogenetic tree of Bigmessidae (clade XVII) from 2011 comprising subclades A to I and the corroborating outgroups XIV to XVI and XVIII to XXI as discerned using combined sequence analyses of nuclear 28S rDNA, histone H3, 5.8S ITS rDNA, and mitochondrial CO1 and IGR loci. All taxa from canonical Faviidae unless associated with symbols where black circles denote definitive clades with ≥98 percent of 1,000 bootstraps, whereas 1, 2, and 3 asterisks represent para-, poly-, or both para- and poly-phyly respectively. Maximum likelihood bootstrap (≥50%)/maximum parsimony bootstrap (≥50%)/Bayesian posterior probability (≥0.9) are associated with each node, while hyphens represent marginal inconsistencies. For scale bar see Appendix I. Tree and analyses courtesy of Huang et al. 2011 and the Creative Commons Attribution License 2.0.
Subclade XVII A comprises two strains of Goniastrea australensis (S088 and S098) and G. favulus (G61877 and S022) and three strains of G. pectinata (G61879, P110, and S043) collected from three Indo-Pacific regions. Subclade XVII B clusters three strains of Favia favus from the Pacific and Singapore, whilst three other strains from GBR and Singapore are clustered with contrasting species. Favia rotundata from GBR and the Philippines is grouped with Favites in subclade XVII F+H which was originally Favites rotundata Veron, Pichon and Wijsman-Best 1977 (Huang et al. 2011). Subclade XVII F is defined by Favites abdita Ellis and Solander 1786-like corals that accommodates Favites Link 1807; Montastraea de Blainville 1830, and Favia Ehrenberg 1834. The former is the oldest radiation and thus Montastraea and Favia of subclade XVII F should be designated Favites, while Favites pentagona, F. russelli and F. stylifera are absorbed by other genera (Fig 19.). Species of Caulastraea, Cyphastrea, Echinopora, Hydnophora, Leptoria, Merulina, and Oulophyllia appear stable, whereas Favites and Platygyra include subclade species architypes, hence these names should endure. Rigorous analyses of Faviidae have identified anomalies akin to Moseleya latistellata and Montastraea multipunctata in Bigmessidae which would be better ranked elsewhere, while the findings of the current study indicated that merely minor revisions are required to establish monophyly (Huang et al. 2011).
Budd and colleagues compiled a review of morphological and molecular techniques used to classify the order Scleractinia which was published in 2010. Given that hard coral classification was founded on the structural features of coral skeletons, it would be remiss to not explore their phylogenetically discerning topologies. The polyps of colonial Anthozoa are supported by a cup called the calice located on the living part of the skeletal surface or corallum. A tubular extension into the corallum is crowned by a corallite upon which most morphological studies rely. All coral accretions are structurally-intricate and -multifaceted while their microporous superficial surfaces are pervaded with living flesh (Budd et al. 2010).

Fig 19. The subclades A to I of Bigmessidae (clade XVII) discerned using combined sequence analyses of nuclear 28S rDNA, histone H3, 5.8S ITS rDNA, and mitochondrial CO1 and IGR loci with emboldened genera-defining archetypes. All taxa from canonical Faviidae unless associated with symbols where A, G, J, P, and S represent strains from the Atlantic, GBR, Japan, the Philippines, and Singapore with suffix GenBank accession numbers. Values above each node express maximum likelihood bootstrap (≥50%)/Bayesian posterior probability (≥0.9). Sub-node integers are maximum parsimony bootstraps (≥50%) while hyphens signify marginal inconsistencies. Tree and analyses courtesy of Huang et al. 2011 and the Creative Commons Attribution License 2.0.
The columella is the corallite’s central axis from which cylindrically arranged dividing septa radiate while their costae are septal extensions that infiltrate the tube-like corallite walls which are merged into a colonial framework by the corallite-adjoining coenosteum or peritheca. Further discerning features include perpendicularly coordinated corallite wall-forming rod-like synapticulae that weave the posterior-septal margin, platform-like diagonal or planar structures (dissepiments) called endotheca and exotheca that lie within and without the corallite wall, whereas epitheca are membrane-like calciferous sheaths that surround corallites or the corallum (Figs 20. & 21.; Budd et al. 2010).
Three kinds of skeletal examinations were categorized by Budd and Stolarski in 2009 that assist taxological elucidation of ancestry: 3D macromorphology where a brightfield microscope is used to scrutinize corallites and their physical context within the colony; micromorphological 3D observations of the features along the septal-facet margins and calical surfaces that focus on dentation and granulation using light or electron microscopy, and 2D microstructural examinations that trace the deposition of perceptible rods called trabeculae within the corallite wall and septa which requires refined sample preparation and a scanning electron microscope (SEM; Budd et al. 2010). Studies of micromorphology facilitate familiar and higher classifications while macromorphology is used for intra-generic and -specific ranks (Wells 1956), and fortuitously, mainstream taxonomy did not rely on microstructural analyses. Septal topology and enumerations of trabeculae combined with corallite wall disparities drove Alloiteau to promote the families Amphiastreidae, Meandrinidae, and Stylinidae to suborders in 1957, while three more were added in 1987 by Chevalier and Beauvais after studying trabeculae formation, consolidating accretions, and the presence or absence of a medioseptal plane (Alloiteau 1957, cited in Budd et al. 2010; Chevalier & Beauvais 1987, cited in Budd et al. 2010).
Early micromorphological analyses focused on septal formation where light microscopy led to their canonical growth paradigm (Ogilvie 1897, cited in Budd et al. 2010; Bryan & Hill 1941, cited in Budd et al. 2010; Vaughan & Wells 1943, cited in Budd et al. 2010; Wells 1956) which proposed that septa were constructed from discrete vertical trabecular rods that terminate in upper marginal teeth that may or may not be fan-like. It was postulated that transverse sections of trabeculae would appear as axes of calcification with radially arranged fibers. Sclerodermite referred to a simple accretion pathway and its fibers, whereas compound trabeculae were bundles or complex configurations of filaments and routes of deposition (Budd et al. 2010). Septal porosity; comparative simple or compound trabeculae per septum; fan-systems per tooth, and synapticulae were considered the most phylogenetically discriminating attributes of the time (Vaughan & Wells 1943, cited in Budd et al. 2010; Wells 1956).
The first five suborders to be defined are featured in table 5. consistent with the historic findings of Vaughan and Wells.

Fig 20. A diagram illustrating the phylogenetically discriminating features of scleractinian skeletons. From the anatomically accurate representation drawn by Wells © 1956 and featured in Budd et al. 2010.
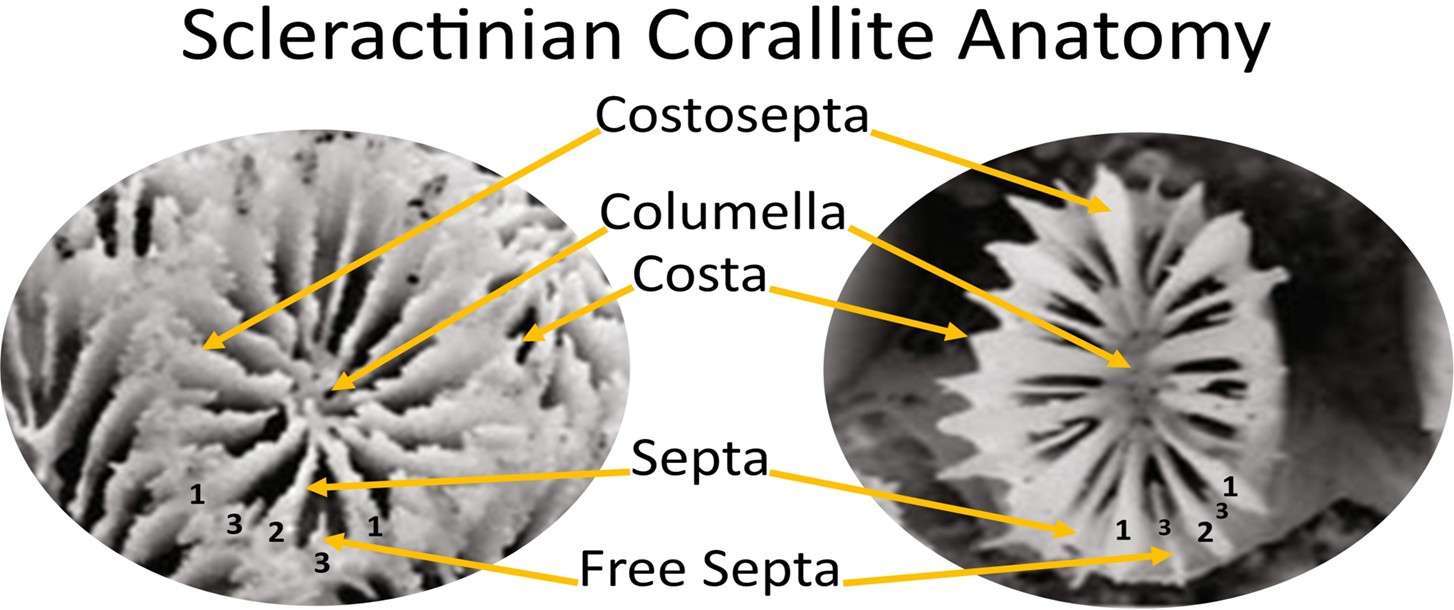
Fig 21. The labelled corallites of Goniastrea aspera (Robusta: Faviidae; left) and Galaxea fascicularis (Complexa: Oculinidae) where Arabic numerals on the costal edge refer to septal cycles. Photography courtesy of Ying et al. 2018 and the Creative Commons Attribution License 4.0.
Micromorphology was also used to rank families where the septal phenotypes of Faviidae, Mussidae, and Merulinidae are described in table 6. (Budd et al. 2010).
Macromorphological analyses evaluate the composition of corallite walls; manifestations of dissepiments, or whether corallites bud from within or from the exterior of calices when the colony grows (Budd et al. 2010).
40 years of technological advancements in electron microscopy have widened our perception of skeletogenesis, insofar as elucidation of independent dual-phase aragonite crystallization and septal formation orchestrated by a fibril-centered organic meshwork (Cuif et al. 1997, cited in Budd et al. 2010; Cuif et al.1999; Cuif & Sorauf 2001, cited in Budd et al. 2010; Cuif et al. 2003; Stolarski 2003; Cuif & Dauphin 2005a; Cuif & Dauphin 2005b). Nevertheless, microstructural analysis was barely established in 2010 (Budd et al. 2010). Axes of accretion form a scaffold that an advancing front sequentially thickens with fibrous deposits from the secretory calicoblastic ectoderm (Budd et al. 2010).

Table 5. The first five scleractinian suborders to be ranked by Vaughan and Wells in the mid-20th century using light microscopy and their discerning septal attributes. Adapted from Budd et al. 2010.
The synchronicity of both skeletogenic phases was first described by Stolarski as simultaneous swift axial concretions and solidifications (Stolarski 2003).
Deposition rapidity, axes, size, shape, magnitudes of thickening, and increments of growth differ amongst taxa. Unsurprisingly, mineral and organic phases are less defined in azooxanthellate Scleractinia compared with hermatypes which perform the planet’s swiftest form of biocalcification, where lipid excretions from their “algal”-partners appear decisive (Cohen & McConnaughey 2003; Munn 2019). The corallum is continuously enlarging along with septal teeth and surface granulations which are best observed using low magnification electron microscopy (Budd et al. 2010). However, powerful stereo microscopy or supplementary high color rendering index (CRI) overhead stage illumination of a brightfield microscope significantly enhances topological acuity.

Table 6. Three scleractinian families classified by Vaughan and Wells in the mid-20th century using light microscopy and their discerning septal attributes. Adapted from Budd et al. 2010.
Table 7. highlights the differences in micromorphology and the microstructural features of Atlantic and Pacific mussids of clades XXI and XIX respectively (Fukami et al. 2008; Budd & Stolarski 2009).
One of the few remarkable findings of morphological studies came from the phenetic work of Powers and Rohlf published in 1972 whose tree was not overly dissimilar to Fukami and collaborators’ molecular reconstruction from 2008, which differentiated the genera Fungia and Psammocora from Complexa and highlighted the polyphyly of Faviidae (Powers & Rohlf 1972; Fukami et al. 2008).
Cairns analysed corallum size and shape, costal disparities, and colonial configuration in addition to the poorly-defined micromorphological traits: thecal porosity, the density and shape of costal spines, and septal tooth dimensions. He recorded spine ornamentation which was not included in his deliberations (Cairns 1984). The same researcher later examined Turbinoliidae and Dendrophylliidae in 1997 and 2001 where he considered the embellishment of costal spines in the former and the synapticulate rods of the latter (Cairns 1997, cited in Budd et al. 2010; Cairns 2001). Hoeksema scrutinized septal perforations and surface granules that extended onto the walls of corallites as well as the structure and enumerations of costal spines and septal teeth. The research unearthed compound synapticulae and five lineages of synapomorphies within Fungiidae (Hoeksema 1989, cited in Budd et al. 2010). The above inquiries emphasize the value of ontogenic biomarkers and attest the efficacy of micromorphological and microstructural analyses which will likely amplify the resolution of molecular techniques (Budd et al. 2010).
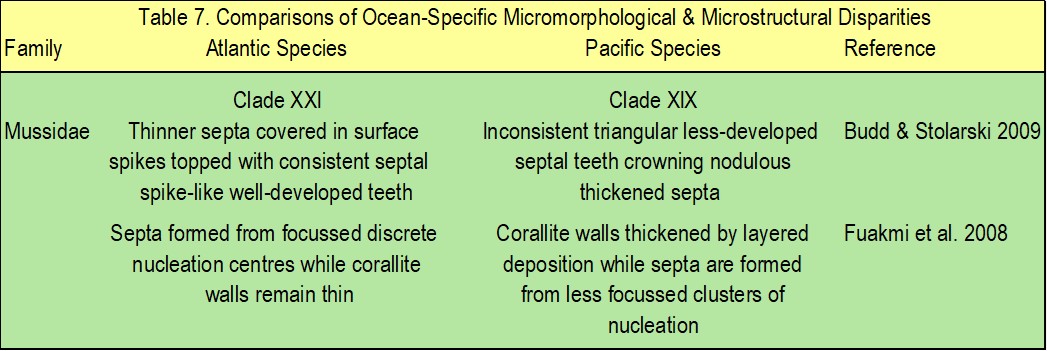
Table 7. The micromorphological and microstructural differences of Atlantic and Pacific mussids of clades XXI and XIX respectively. Adapted from Budd et al. 2010.

Fig 22. The monophyletic tree of Ramirez-Portilla and allies from 2022 who used bioinformatics, analytical software, sequencing, and morphological traits to delineate Japan’s clades of tabular Acropora cytherea, A. hyacinthus, and A. bifurcata that can survive at mesophotic depth (Fig 23.). Adapted from Ramirez-Portilla et al. 2022
The divergence of Robusta and Complexa means that the subordinal classifications ranked and reported by Wells in 1956 are polyphyletic, while Dendrophylliidae is conspicuous amongst the 10 remaining extant monophyletic families. These clades now comprise corals that were formerly ranked elsewhere, whereas some genera are also para- or poly-phyletic. More uncertainty arises from the recognition of a Basal clade and the polyphyly of the deep-water azooxanthellate family Caryophylliidae (Budd et al. 2010; Kitahara et al. 2010). However, these are all signs that we have a definitive means of determining the origins of species, while molecular analyses are limited insofar as nucleic acids cannot be extracted from fossils. Notwithstanding the ostensibly foreseen unrivalled resolution offered by mitochondrial sequence analyses, such genes are highly conserved, and thus scarcely diverge. Hence discernment of higher classifications proves challenging when mitotyping using ribosome- or respiratory chain component-encoding genes. Most hermatypic corals broadcast both kinds of gametes which is their original procreative strategy that increases the likelihood of interspecific hybridisation (Baird et al. 2009; Budd et al. 2010). Interbreeding of Acropora cf. bifurcata, A. cf. cytherea, and A. cf. hyacinthus like most interspecific crosses led to reproductive failure (Ramirez-Portilla et al. 2022) likely due to non-fertilization or chromosome instability (Oliver & Babcock1992; Willis et al. 1997; Huret et al. 2000). However, it did affirm them as distinct species, while often precise conspecific broadcasting windows (Levitan et al. 2004; Baird et al. 2009) and temporal meiocyte viability (Levitan et al. 2004) makes outcrosses unlikely. Even so, the findings of contrasting research were redolent of frequent hybridization (Vize et al. 2005; Levitan et al. 2011) while Acropora prolifera remains a stable cross of A. cervicornis and A. palmata (Pinzón et al. 2010; Reich et al. 2021).
Furthermore, allorecognition effectors are implicated in the detection and inhibition of chimeras (Miller et al. 2007) while conspecific spats fuse which enhances performance, although tissue rejection occurs along colony boundaries when they mature (Raymundo & Maypa 2004). Contrary to the species of Acropora cannot be phylogenetically-discerned due to interspecific hybridization paradigm, they demonstrate extraordinary fidelity, while their lineages evolve independently (Fig 22.; Ramirez-Portilla et al. 2022). Nevertheless, protracted introgressive hybridization whereby genetic material is bidirectionally shared between species, creates viable and fertile crosses even amongst reef teleosts (Combosch & Vollmer 2015; Suzuki et al. 2016; Harrison et al. 2017).

Fig 23. A mesophotic-dwelling table-like (tabular) colony of Acropora cytherea susceptible to bleaching caused by strains of the bacterium Vibrio coralliilyticus (Sussman et al. 2008).
Reverse taxological molecular studies will conceivably pinpoint definitive novel anatomies which may be traced through the fossil record to the present day, where combining historic inquiries of numerous skeletal features onto a single tree will assist formulation of hypotheses for seminal gene sequence analyses. Macromorphological examinations of epitheca suggest that they were lacking more than once on evolutionary timescales in clades XVII, XIX, and XXI where the former reverted, whereas elliptical septal teeth became circular much earlier than epithecal loss in clades XVI/XVII, XIX, and XXI where they re-evolved, hence these features appear more developmentally-plastic. Although rigorous data analyses were redolent of insignificant disparities between the rates of gain and loss, epitheca have vanished six times and reemerged merely twice. Nevertheless, these findings demonstrate that phylogenetic-competency is commensurate with a broadening of morphological datasets and bioinformatics (Budd et al. 2010).
All the same, several questions remain unanswered. Do a handful of definitive phylogenetically-discerning skeletal topologies exist. Are some coralla predisposed to rapid change. Do these changes have trajectory bias. And are some morphological traits linked to rates of diversification (Budd et al. 2010).
That concludes phase 1. of this academic series while next we explore further revisions arising from the establishment of familiar and tentative ordinal monophyly to explain the reclassifications in Fukami and Kitano from 2023.

Fig 24. A phylogenetic tree (cladogram) representing the evolutionary histories of 121 Scleractinia discerned using Bayesian analyses of concatenated mitochondrial 16S rDNA and nuclear 28S rDNA with the corroborating outgroup Ricordea florida (Corallimorpharia) where families are color coded. Species that were molecularly analyzed in this study are emboldened, whereas coralla that epitomize each genera are illustrated to the right. Featuring the early divergence of the Basal clade represented by the families Gardineriidae and Micrabaciidae and the later Complexa and Robusta split. [*] maximum likelihood Chi-square >0.95 and bootstrap >70% with posterior probability >90; [+] maximum likelihood Chi-square >0.8 and bootstrap >55% with posterior probability >80; [ ■] zooxanthellate; [●] colonial. The study’s investigations of rDNA markers and the fossil record maintained that Scleractinia evolved from a Palaeozoic soft-bodied ancestor distinct from the order Rugosa whose skeletons were calcite and not aragonite. Several Permian scleractiniamorphs like Houchangocyathus wangi and H. yaoi that perished after the mass extinction constructed aragonite skeletons, whereas mid-Ordovician Kilbuchophyllum discoidea (Kilbuchophyllida: Kilbuchophyllidae) had corallite walls like countless modern stony corals (Scrutton & Clarkson 1991; Scrutton 1993; Ezaki 1997; Ezaki 2000; Romano & Cairns 2000). Adapted from and courtesy of Stolarski et al. 2011 and the Creative Commons Attribution Licence.
Allele refers to different forms of a gene.
Animal Pole is a surface zone of an egg which marks the most mitotically active yolk-diminished region.
Bootstrap replicates are equal-sized multiple sequence aligned fictional substitute nucleic acids with confidences proportional to percentage identity.
Canonical, substantiated, acknowledged.
Clade describes a monophyletic group of organisms comprising all the descendants from a common ancestor (Cladistics).
Costa is a radial structure outside the corallite wall which may be autonomous or integral to a costoseptum.
Costoseptum is a fan-shaped radial element that adjoins an outer costa to a conspicuous inner septum unified with the upper columella (Budd & Stolarski 2011).
Congeners refers to intrageneric relatives (Congeneric).
The cells of a Diploid organism have a chromosome from each parent (2n), whereas the copy number of Haploid organisms is 1n.
Fenestrate refers to perforations or transparencies.
Hermatypic and Ahermatypic Corals – the terms hermatypic/ahermatypic have been misused because irregularities such as sea fans possessing rigid skeletons are constructive, yet they may or may not contain “algal” symbionts and do not contribute to reef substratum. Ahermatypes may have large or small polyps and do not contain zooxanthellae. Scleractinian corals are “true” reef builders which may nurture “algal” partners or be azooxanthellate. The terms Scleractinia and hermatype are not interchangeable, inasmuch as hermatypic corals contain zooxanthellae and are constructive, whereas ahermatypes are azooxanthellate and non-constructive. This clear definition also applies to non-Scleractinia (Schuhmacher & Zibrowius 1985).
Heterotrophs cannot utilize inorganic carbon for growth, so they must dine on organic plant or animal derived nourishment. Their sources include glucose, yet several microscopic taxa retain the capacity to transform inorganic compounds which are frequently used as substrates for chemical reactions.
Heterozygous refers to different forms of a gene carried by a diploid (2n) organism usually from each parent on opposing chromosomes where each one of an individual’s 1n gametes carry one kind of allele.
Homoplasy and Convergent Evolution are synonymous insofar as they refer to the acquisition of similar or identical traits in unrelated organisms.
Homozygous refers to the same form of a gene carried by a diploid (2n) organism usually from each parent on opposing chromosomes where each one of an individual’s 1n gametes carry the same kind.
Interspecific Hybridization creates a subpopulation with a distinct genotype.
Introgression facilitates the dissemination of genes between species arising from recurring hybridization and backcrossing over long periods of time.
Meiocyte is a cell generated by meiosis which is contextually synonymous with Gamete within which diploid chromosome copy number is halved (Haploid).
Ontogeny/Ontogenesis describes the development of an organism from the moment of fertilization to a mature adult.
Ontology is used contextually to describe the intra- and extra-cellular, ana-, cata-, and meta-bolic pathways associated with an organism’s day-to-day housekeeping, homeostatic, and reproductive processes.
Paralogues are two or more copies derived from a single ancestral gene residing at multiple distal loci in the same genome.
Paraphyletic refers to a group of organisms that evolved from a common ancestor that lacks all descendants.
The corallites of Phaceloid skeletons form distinct branches which lack coenosteum.
Phenetics, Numerical Taxonomy, or Taximetrics classify organisms according to their physical features irrespective of their putative evolutionary trajectories.
Phylogenetic Tree “is a graphic representation of the interrelations and evolutionary history of a group of organisms, indicating the relative order of successive divisions of the line of descent, coincident with speciation events” (Wallace et al. 1996).
Synonymous Plesiomorphy and Symplesiomorphy refer to the sharing of an inherited attribute by all members of a clade which does not differentiate that clade from others.
The Scale Bars of Phylogenetic Trees refer to relative evolutionary distance expressed as the developmental substitutions of nucleotides.
Svedberg (S) is a sedimentation coefficient that relates to particle-size and settlement-rapidity during centrifugation. 1 svedberg is equal to 10-13 seconds (100 femtoseconds) which is a non-SI unit used to describe the size of nucleic acids or proteins (Stryer 1995a).
Symbiosis is the coexistence of two or more closely associated organisms which acquire mutual benefit.
Synapomorphies are modified forms of an ancestral attribute that are exclusively shared amongst all descendants. Or, Synapomorphy refers to phylogenetically-discriminating empirical attributes that epitomize a monophyletic clade (Hennig 1957, cited in Arrigoni et al. 2019).
Synapticulae are perpendicularly-coordinated corallite wall-forming rods that interweave the posterio-septal margin.
Syntenic means “threaded together” which refers to several intrachromosomal genes traced using somatic cell hybridization (Russell 1998).
Trabeculae were thought septum-forming discrete vertical rods associated with radially arranged fibers while they are now known to be smooth centers of rapid aragonite deposition that craft numerous skeletal features (Nothdurft & Webb 2007).
The terms Zooxanthellate and Azooxanthellate refer to organisms with and without Symbiotic “algal”-partners.
References
Adams, J. (2018) Chalice Coral Family Gets New Paraechinophyllia Genus. ReefBuilders.com. https://reefbuilders.com/2018/08/17/chalice-coral-family-gets-new-paraechinophyllia-genus/
Allen, J., F. (1993) Control of gene-expression by redox potential and the requirement for chloroplast and mitochondrial genomes. J. Theor. Biol. 165, 609-631. http://www.doi.org/10.1006/jtbi.1993.1210
Allen, J., F. (2003) The function of genomes in bioenergetic organelles. Phil. Trans. R. Soc. Lond. B 358, 19-37. https://www.doi.org/10.1098/rstb.2002.1191
Alloiteau, J. (1957) Contribution a la syste´matique des madre´- poraires fossiles. Paris: Centre National de la Recherche Scientifique. Washington (DC): Smithsonian Institution Press. pp 1–55.
Arrigoni, R., Benzoni, F., Huang, D., Fukami, H., Chen, C., A., Berumen, M., L., Hoogenboom, M., Thomson, D., P., Hoeksema, B., W., Budd, A., F., Zayasu, Y., Terraneo, T., I., Kitano, Y., F. & Baird, A., H. (2016) When forms meet genes: revision of the scleractinian genera Micromussa and Homophyllia (Lobophylliidae) with a description of two new species and one new genus. Contributions to Zoology. 85, 387-422.
Arrigoni, R., Berumen, M., L., Stolarski, J., Terraneo, T., I. & Benzoni, F. (2019) Uncovering hidden coral diversity: a new cryptic lobophylliid scleractinian from the Indian Ocean. Cladistics. 35, 301-328.
Arrigoni, R., Huang, D., Berumen, M., L., Budd, A., F., Montano, S., Richards, Z., T., Terraneo, T., I. & Benzoni, F. (2021) Integrative systematics of the scleractinian coral genera Caulastraea, Erythrastrea and Oulophyllia. Zoologica Scripta. 50, 509-527.
Arrigoni, R., Kitano, Y., F., Stolarski, J., Hoeksema, B., W., Fukami, H., Stefani, F., Galli, P., Montano, S., Castoldi, E. & Benzoni, F. (2014) A phylogeny reconstruction of the Dendrophylliidae (Cnidaria, Scleractinia) based on molecular and micromorphological criteria, and its ecological implications. Zoologica Scripta. 43, 661-688.
Arrigoni, R., Stolarski, J., Terraneo, T., I., Hoeksema, B., W., Berumen, M., L., Payri, C., Montano, S. & Benzoni, F. (2023) Phylogenetics and taxonomy of the scleractinian coral family Euphylliidae. Contributions to Zoology. 92, 130-171
Aslett, C., G. (2024) The Complete Reef Aquarist, for hobbyists, the trade and academics – A Conservation Manual. Aslett, C., G. (ed.). Reef Ranch Publishing Ltd, Leeds, West Yorkshire, UK. https://www.ebay.co.uk/itm/116390701502 pp 402-407.
Baird, A., H., Guest, J., R. & Willis, B., L. (2009) Systematic and biogeographical patterns in the reproductive biology of scleractinian corals. Annual Review of Ecology, Evolution, and Systematics. 40(1), 551-557. http://www.dio.org/10.1146/annurev.ecolsys.110308.120220
Beckmann, A. & Özbek, S. (2012) The nematocyst: a molecular map of the cnidarian stinging organelle. The International journal of developmental biology. 56(6-8), 577-582.
Bonacolta, A., M., Weiler, B., A., Porta‐Fitó, T., Sweet, M., Keeling, P. & del Campo, J. (2023) Beyond the Symbiodiniaceae: diversity and role of microeukaryotic coral symbionts. Coral Reefs. 42, 567-577. http://doi.org/10.1007/s00338-023-02352-0
Bryan, W., H. & Hill, D. (1941) Spherulitic crystallization as a mechanism of skeletal growth in the hexacorals. Proc Royal Soc Queensland, 52, 78-91.
Budd, A., F. & Stolarski, J: (2009) Searching for new morphological characters in the systematics of scleractinian reef corals: comparison of septal teeth and granules between Atlantic and Pacific Mussidae. Acta Zool. 90, 142-165.
Budd, A., F. & Stolarski, J: (2011) Corallite wall and septal microstructure in scleractinian reef corals: Comparison of molecular clades within the family Faviidae. J Morphol. 272, 66-88.
Budd, A., F., Romano, S., L., Smith, N., D. & Barbeitos, M., S. (2010) Rethinking the phylogeny of scleractinian corals: A review of morphological and molecular data. Integr Comp Biol. 50, 411-427. http://www.doi.org/10.1093/icb/icq062
Cairns, S., D. (1984) An application of phylogenetic analysis to the Scleractinia: family Fungiidae. Palaeontographica Americana. 54, 49-57.
Cairns, S., D. (1997) A generic revision and phylogenetic analysis of the Turbinoliidae (Cnidaria: Scleractinia). Smithsonian Contributions to Zoology, Vol. 591.
Cairns, S., D. (2001) A generic revision and phylogenetic analysis of the Dendrophylliidae (Cnidaria: Scleractinia). Smithson Contrib Zool. 615, 75.
Cavalier-Smith, T. & Chao, E., E., Y. (2006) Phylogeny and Megasystematics of Phagotrophic Heterokonts (Kingdom Chromista). J Mol Evol. 62, 388-420.
Cavalier-Smith, T. (2010) Kingdoms Protozoa and Chromista and the meozoan root of the eukaryotic tree. Biol. Lett. 6, 342-345.
Cavalier-Smith, T. (2018) Kingdom Chromista and its eight phyla: a new synthesis emphasising periplastid protein targeting, cytoskeletal and periplastid evolution, and ancient divergences. Protoplasma. 255, 297-357.
Chen, C., A., Odorico, D., M., Ten Louis, M., Veron, J., E., N. & Miller, D., J. (1995) Systematic relationships within the Anthozoa (Cnidaria Anthozoa) using the 59-end of the 28S rDNA. Mol Phylogenet Evol. 4(2), 175-183.
Chevalier, J-P. & Beauvais, L. (1987) Ordre de Scle´ractiniaires. In: Grasse PP, editor. Traite´ de Zoologies, Cnidaires, Anthozoaires. Paris: Masson. pp 403-764.
Cohen, A. & McConnaughey, T., A. (2003) Geochemical Perspectives on Coral Mineralization. Reviews in Mineralogy & Geochemistry. 54, 151-187.
Combosch, D., J., Vollmer, S., V. (2015) Trans-Pacific RAD-Seq population genomics confirms introgressive hybridization in Eastern Pacific Pocillopora corals. Mol. Phylogenet. Evol. 88, 154–162.
Cuif, J., P., Lecointre, G., Perrin, C., Tillier, A. & Tillier. S. (2003) Patterns of septal biomineralization in Scleractinia compared with their 28S rDNA phylogeny: a dual approach for a new taxonomic framework. Zool Scripta. 32, 459-473.
Cuif, J-P. & Dauphin, Y. (2005a) The two-step mode of growth in the scleractinian coral skeletons from the micrometer to the overall scale. J Struct Biol. 150, 319-31.
Cuif, J-P. & Dauphin, Y. (2005b) The environmental recording unit in coral skeletons – a synthesis of chemical and structural evidences for a biochemically driven, stepping-growth process in fibers. Biogeosciences. 2, 61-73.
Cuif, J-P. & Sorauf, J., E. (2001) Biomineralization and diagenesis in Scleractinia: part I, biomineralization. Bull Tohoku Univ Museum. 1, 144-151.
Cuif, J-P., Dauphin, Y. & Gautret, P. (1997) Biomineralization features in scleractinians coral skeletons: source of new taxonomic criteria. Boletın Real Sociedad Espanola de Historia Natural, Seccio´n Geologica. 92,129-141.
Cuif, J-P., Dauphin, Y. & Gautret, P. (1999) Compositional diversity of soluble mineralizing matrices in some recent coral skeletons compared to fine-scale growth structures of fibers: discussion of consequences for biomineralization and diagenesis. Int J Earth Sci. 88, 582-92.
Cuif, J-P., Lecointre, G., Perrin, C., Tillier, A. & Tillier, S. (2003) Patterns of septal biomineralization in Scleractinia compared with their 28S rRNA phylogeny: a dual approach for a new taxonomic framework. Zool Scripta. 32, 459-473.
Daly, M., Brugler, M., Cartwright, P., Collins, A., Dawson, M., Fautin, D., France, S., McFadden, C., Opresko, D., Rodriguez, E., Romano, S. & Stake, J. (2006) The Phylum Cnidaria: A Review of Phylogenetic Patterns and Diversity 300 Years After Linnaeus. Zootaxa. 1668,.
Ezaki, Y (1997) The Permian coral Numidiaphyllum: new insights into anthozoan phylogeny and Triassic scleractinian origins. Palaeontology. 40, 1-14.
Ezaki, Y. (2000) Palaeoecological and phylogenetic implications of a new scleractiniamorph genus from Permian sponge reefs, south China. Palaeontology. 43, 199-217.
Fabricius, K. (2011) Octocorallia. Encyclopaedia of Modern Coral Reefs. Hopley, D. (ed.). Springer, Netherlands. pp 740-745.
Filkorn, H., F. & Allor, J., P. (2004) A new early cretaceous coral (Anthozoa; Scleractinia; Dendrophylliina) and its evolutionary significance. J Paleont. 78(3), 501-512.
Fukami, H. & Kitano, Y., K. (2023) Changes of scientific names of the scleractinian corals until 2023. Journal of the Japanese Coral Reef Society. 25, 19-25.
Fukami, H. & Nomura, K: (2009) Existence of a cryptic species of Montastraea valenciennesi (Milne Edwards and Haime, 1848) in Wakayama, southern Honshu, Japan (in Japanese). J Jpn Coral Reef Soc. 11, 25-31.
Fukami, H., Chen, C., A., Budd, A., F., Collins, A., Wallace, C. et al. (2008) Mitochondrial and nuclear genes suggest that stony corals are monophyletic but most families of stony corals are not (Order Scleractinia, Class Anthozoa, Phylum Cnidaria). PLOS ONE. 3.
Gong, S., Liang, J., Jin, X., Xu, L., Zhao, M. & Yu, K. (2023) Unfolding the secrets of microbiome (Symbiodiniaceae and bacteria) in cold-water coral. Environmental Microbiology. 11(5). https://doi.org/10.1128/spectrum.01315-23
Harrison, H., B., Berumen, M., L., Saenz-Agudelo, P., Salas, E., Williamson, D., H. & Jones, G., P. (2017) Widespread hybridization and bidirectional introgression in sympatric species of coral reef fish. https://doi.org/10.1111/mec.14279
Hennig, W. (1957) Systematik und Phylogenese. In: Hannemann, H.-J. (Ed.), Berichtuber die Hundertjahrfeier der Deutschen Entomologischen Gesellschaft Berlin. Akademie Verlag, Berlin. pp 50-71.
Hodgson G: (1985) A new species of Montastrea (Cnidaria, Scleractinia) from the Philippines. Pac Sci. 39, 283-290.
Hoeksema, B., W. (1989) Taxonomy, biogeography, and phylogeny of mushroom corals (Scleractinia: Fungiidae). Zoologische Verhandelingen Leiden. 254, 295.
Huang, D., Arrigoni, R., Benzoni, F., Fukami, H., Knowlton, N., Smith, N., D., Stolarski, J., Chou, L., M. & Budd, A., F. (2016) Taxonomic classification of the reef coral family Lobophylliidae (Cnidaria: Anthozoa: Scleractinia). Zoological Journal of the Linnean Society. 178, 436-481.
Huang, D., Meier, R., Todd, P., A. & Chou, L., M: (2009) More evidence for pervasive paraphyly in scleractinian corals: Systematic study of Southeast Asian Faviidae (Cnidaria; Scleractinia) based on molecular and morphological data. Mol Phylogenet Evol. 50, 102-116.
Huang, D., W., Licuanan, W., Y., Baird, A., H. & Fukami, H. (2011) Cleaning up the ‘Bigmessidae’: Molecular phylogeny of scleractinian corals from Faviidae, Merulinidae, Pectiniidae and Trachyphylliidae. BMC Evol Biol. 11, 37.
Huret, J., L., Leonard, C. & Savage, J., R., K. (2000) Chromosomes, Chromosome Anomalies. MRC Radiation, Genome Stability Unit, Harwell, Didcot, OX11 0RD, UK. https://atlasgeneticsoncology.org/teaching/30084/chromosomes-chromosome-anomalies
Janouškovec, J., Horak, A., Obornik, M., Lukes, J. & Keeling, P., J. (2010) A common red algal origin of the apicomplexan, dinoflagellate, and heterokont plastids. Proc Natl Acad Sci. 107, 10949-10954.
Kayal, E., Roure, B., Philippe, H., Collins, A., G. & Lavrov, D., V. (2013) Cnidarian phylogenetic relationships as revealed by mitogenomics. BMC Evol Biol. 13, 5.
Kerr, A., M., Baird, A., H. & Hughes, T., P: (2011) Correlated evolution of sex and reproductive mode in corals (Anthozoa: Scleractinia). Proc R Soc B-Biol Sci. 278, 75-81.
Kitahara, M., V., Cairns, S., D., Stolarski, J., Blair, D. & Miller, D., J. (2010) A comprehensive phylogenetic analysis of the Scleractinia (Cnidaria, Anthozoa) based on mitochondrial CO1 sequence data. PLOS ONE. 5.
Kitahara, M., V., Fukami, H., Benzoni, F. & Huang, D. (2016) The new systematics of Scleractinia: integrating molecular and morphological evidence. The Cnidaria, past, present and future. Goffredo, S. & Dubinsky, Z. (eds.). Springer, Cham. pp 41-59.
Kooistra, W., Gersonde, R., Medlin, L. & Mann, D. (2007) Chapter 11, The Origin and Evolution of the Diatoms. Their Adaptation to a Planktonic Existence. Evolution of Primary Producers in the Sea. Falkowski, P., G. & Knoll, A., H. (eds.). Academic Press, Burlington. pp 207-249.
LaJeunesse, T., C., Parkinson, J., E., Gabrielson, P., W., Jeong, H., J., Reimer, J., D., Voolstra, C., R. & Santos, S., R. (2018) Systematic Revision of Symbiodiniaceae Highlights the Antiquity and Diversity of Coral Endosymbionts. Current biology: CB. 28(16), 2570-2580.
Levitan, D., R., Fogarty, N., D., Jara, J., Lotterhos, K., E. & Knowlton, N. (2011) Genetic, spatial, and temporal components of precise spawning synchrony in reef building corals of the Montastraea annularis species complex. Evolution. 65, 1254-1270.
Levitan, D., R., Fukami, H., Jara, J., Kline, D., McGovern, T., M., McGhee, K., E., Swanson, C., A. & Knowlton, N. (2004) Mechanisms of Reproductive Isolation Among Sympatric Broadcast-Spawning Corals of Montastraea annularis species complex. Evolution. 58, 308-323.
Löser, H., Angel Fernández-Mendiola, P., Pérez-Malo, J., Domínguez Pascual, S. & Cahuzac, B. (2021) Redefinition of the family Rhizangiidae (Scleractinia; Cretaceous to Recent) and description of a new genus from the Early Cretaceous of Spain. Neues Jahrbuch für Geologie und Paläontologie – Abhandlungen. 299, 259-274.
Luzon, K., S., Lin, M., F., Ablan Lagman, M., C., A., Licuanan, W., R., Y. & Chen, C., A. (2017) Resurrecting a subgenus to genus: molecular phylogeny of Euphyllia and Fimbriaphyllia (order Scleractinia; family Euphyllidae; clade V). PeerJ. 5, e4074.
Maier, U-G., Zauner, S., Woehle, C., Bolte, K., Hempel, F., Allen, J., F. & Martin, W., F. (2013) Massively convergent evolution for ribosomal protein gene content in plastid and mitochondrial genomes. Genome Biol. Evol. 5, 2318-2329. http://www.doi.org/10.1093/gbe/evt181
Malakhov, V., V. (2016) Symmetry and the tentacular apparatus in Cnidaria. Russ J Mar Biol. 42, 287-298.
Manktelow, M. (2010) Lecture: History of Taxonomy. Dept of Systematic Biology, Evolutionary Biology Centre, Uppsala University. http://www.atbi.eu/summerschool/files/summerschool/Manktelow_Syllabus.pdf
Martin, W., F., Garg, S. & Zimorski, V. (2015) Endosymbiotic theories for eukaryote origin. Phil. Trans. R. Soc. B 370, 20140330. http://dx.doi.org/10.1098/rstb.2014.0330
Medina, M., Collins, A., G., Takaoka, T., L., Kuehl, J., V. & Boore, J., L. (2006) Naked corals: skeleton loss in Scleractinia. PNAS. 103(24), 96-100.
Mereschkowsky, C. (1905) U¨ber Natur und Ursprung der Chromatophoren im Pflanzenreiche. Biol. Centralbl. 25, 593-604.
Miller, D., J., Hemmrich, G., Ball, E., E. et al. (2007) The innate immune repertoire in Cnidaria – ancestral complexity and stochastic gene loss. Genome Biology. 8, R59.
Morice, A. (2022) A Complete Guide For Chalice Coral Care and Propagation. BestFishKeeping.com. https://www.bestfishkeeping.com/chalice-coral/#h-what-are-chalice-coral
Munn, C., B. (2019) Marine Microbiology: Ecology & Applications, Third Edition. Munn, C., B. (ed.). CRC Press, Taylor & Francis Group, London. pp 273-326.
Nothdurft, L. D. & ·Webb, G., E. (2007) Microstructure of common reef-building coral genera Acropora, Pocillopora, Goniastrea and Porites: constraints on spatial resolution in geochemical sampling. Facies. 53, 1-26. http://www.dio.org/10.1007/s10347-006-0090-0
Ogilvie, M., M. (1897) Die Korallen der Strambergerschichten. Palaeontographica (Suppl). 2, 73-282.
Okubo, N., Mezaki, T., Nozawa, Y., Nakano, Y., Lien, Y-T., et al. (2013) Comparative Embryology of Eleven Species of Stony Corals (Scleractinia). PLoS ONE 8(12), e84115. http://www.doi.org/10.1371/journal.pone.0084115
Pinzón, J., Devlin-Durante, M., Weber, M., Baums, I. & LaJeunesse, T. (2010) Microsatellite loci for Symbiodinium A3 (S. fitti) a common algal symbiont among Caribbean Acropora (stony corals) and Indo-Pacific giant clams (Tridacna). Conservation Genetics Resources. 3(1), 45-47.
Powers, D., A. & Rohlf, F., J. (1972) A numerical taxonomic study of Caribbean and Hawaiian reef corals. Syst Zool. 21, 53-64.
Pray, L. (2008) Discovery of DNA structure and function: Watson and Crick. Nature Education 1(1), 100.
Quek, Z., B., R., Jain, S., S., Richards., Z., T., Arrigoni, R., Benzoni, F., Hoeksema, B., W., Carvajal, J., I., Wilson, N., G., Baird, A., H., Kitahara, M., V., Seiblitz, I., G., Vaga, C., F. & Huang, D. (2023) A hybrid-capture approach to reconstruct the phylogeny of Scleractinia (Cnidaria: Hexacorallia). Molecular Phylogenetics and Evolution. 186, 107867.
Ramirez-Portilla, C., Baird, A., H., Cowman3, P., F., Quattrini, A., M., Harii, S., Frederic Sinniger, F. & Flot, J-F. (2022) Solving the Coral Species Delimitation Conundrum. Syst. Biol. 71(2), 461-475. http://www.dio.org/10.1093/sysbio/syab077
Raymundo, L., J. & Maypa, A., P. (2004) Getting bigger faster: mediation of size-specific mortality via fusion in juvenile coral transplants. Ecological Applications. 14, 281-295.
Reich, H., Kitchen, S., Stankiewicz, K., Devlin‐Durante, M., Fogarty, N. & Baums, I. (2021) Genomic variation of an endosymbiotic dinoflagellate (Symbiodinium ‘fitti’) among closely related coral hosts. Molecular Ecology. 30(14), 3500-3514.
Romano, S., L. & Cairns, S., D. (2000) Molecular phylogenetic hypotheses for the evolution of scleractinian corals. Bull Mar Sci. 67(3), 1043-1068.
Romano, S., L. & Palumbi, S., R. (1996) Evolution of scleractinian corals inferred from molecular systematics. Science. 271. 640-642. http://www.doi.org/10.1126/science.271.5249.640
Rowlett, J. (2020) Indo-Pacific Corals. Rowlett (self-published). pp 1-809.
Russell, P., J. (1998) Genetics Fifth Edition. Russell, P., J., Buck, E & Dhillon, A. (eds.). The Benjamin/Cumming Publishing Company Inc., Melo Park, CA, USA. p 183; G 8.
Russell, P., J. (1998a) Genetics Fifth Edition. Russell, P., J., Buck, E & Dhillon, A. (eds.). The Benjamin/Cumming Publishing Company Inc., Melo Park, CA, USA. p 694.
Schuhmacher, H. & Zibrowius, H. (1985) What is hermatypic? Coral Reefs. 4, 1-9.
Scrutton, C., T. & Clarkson, E., N., K. (1991) A new scleractinian-like coral from the Ordovician of the Southern Uplands, Scotland. Palaeontology. 34, 179-194.
Scrutton, C., T. (1993) A new kilbuchophyllid coral from the Ordovician of the Southern Uplands, Scotland. Cour Forsch Senck 1993, 164:153-158.
Stanley, Jr. G., D. & Swart, P., K. (1995) Evolution of the coral-zooxanthellae symbiosis during the Triassic: A geochemical approach. Paleobiol. 21, 179-199.
Stolarski, J. & Vertino, A: (2007) First Mesozoic record of the scleractinian Madrepora from the Maastrichtian siliceous limestones of Poland. Facies. 53:67-78.
Stolarski, J. (2003) Three-dimensional micro- and nanostructural characteristics of the scleractinian coral skeleton: a biocalcification proxy. Acta Paleontol Polo. 48, 497–530.
Stolarski, J., Kitahara, M., V, Miller, D., J., Cairns, S., D., Mazur, M. & Meibom, A. (2011) The ancient evolutionary origins of Scleractinia revealed by azooxanthellate corals. BMC Evolutionary Biology. 11, 316.
Stryer, L. (1995a) Biochemistry 4th Edition. Stryer, L. (ed.). W. H. Freeman and Company, New York. pp 653-682.
Sussman, M., Willis, B., L., Victor, S. & Bourne, D., G. (2008) Coral pathogens identified for White Syndrome (WS) epizootics in the Indo-Pacific. PLoS ONE. 3(6), e2393.
Suzuki, G., Keshavmurthy, S., Hayashibara, T., Wallace, C., C., Shirayama, Y., Chen, C., A., Fukami, H. (2016) Genetic evidence of peripheral isolation and low diversity in marginal populations of the Acropora hyacinthus complex. Coral Reefs. 35, 1419–1432.
Technau, U. & Steele, R., E. (2011) Evolutionary crossroads in developmental biology: Cnidaria. Development (Cambridge, England). 138(8), 1447-1458.
Vaughan, T., W. & Wells, J., W. (1943) Revision of the suborders, families and genera of the Scleractinia. Geological Society of America Special Papers Vol. 44. New York: Geological Society of America.
Veron, J., E., N. (1995) Corals in Space and Time: the Biogeography and Evolution of the Scleractinia. Ithaca: Comstock/Cornell paperbacks, Cornell University Press.
Veron, J., E., N., Odorico, D., M., Chen, C., A. & Miller, D., J. (1996) Reassessing evolutionary relationships of scleractinian corals. Coral Reefs. 15, 1-9.
Veron, J., E., N., Pichon, M. & Wijsman-Best, M: (1977) Scleractinia of Eastern Australia. Part II. Families Faviidae, Trachyphylliidae. Australian Institute of Marine Science Monograph Series. 1-233.
Veron, J., E., N: (1986) Corals of Australia and the Indo-Pacific Townsville: Australian Institute of Marine Science.
Vize, P. D., Embesi, J., A., Nickell, M., Brown, D., P & Hagman, D., K. (2005) Tight temporal consistency of coral mass spawning at the Flower Garden Banks, Gulf of Mexico, from 1997-2003. Gulf Mex. Sci. 23, 107-114.
Waggoner, B., M., Collins, A. & Smith D. (1994) Introduction to the Scleractinia. Berkeley University of California Museum of Palaeontology. UCMP.Berkeley.edu. https://ucmp.berkeley.edu/cnidaria/scleractinia.html
Wallace, C., C., Chen, C., A., C., Fukami, H. & Muir, P., R. (2007) Recognition of separate genera within Acropora based on new morphological, reproductive, genetic evidence from A. togianensis, elevation of the subgenus Isopora Studer, 1878 to genus (Scleractinia: Astrocoeniidae; Acroporidae). Coral Reefs 26, 231-239.
Wallace, R., A., Sanders, G., P. & Ferl, R., J. (1996) Biology: The Science of Life, Fourth Edition, HaperCollins Publishers Inc. New York, USA. G-22; pp 384-385.
Watson, J., D. & Crick, F., H., C., A. (1953) structure for deoxyribose nucleic acid. Nature. 171, 737-738.
Wells, J., W. (1956) Scleractinia. Treatise on invertebrate paleontology: coelenterata. Moore, R., C. (ed.). Lawrence: Geological Society of America and University of Kansas Press. pp 328-443.
Willis, B., L., Babcock, R., C., Harrison, P., L. & Wallace, C., C. (1997) Experimental hybridisation and breeding incompatibilities within the mating systems of mass spawning reef corals. Coral Reefs. 16, S53-S65.
WoRMS (2020) World Register of Marine Species. Acropora millepora (Ehrenberg, 1834). http://www.marinespecies.org/aphia.php?p=taxdetails&id=207023
Ying, H., Cooke, I., Sprungala, S., Wang, W., Hayward, D., C., Tang, Y.; Huttley, G., Ball, E., E., Forêt, S. & Miller, D., J. (2018) Comparative genomics reveals the distinct evolutionary trajectories of the robust and complex coral lineages. Genome Biology. 19, 175. http://doi.org/10.1186/s13059-018-1552-8
Disclaimer – every effort has been made to adhere to copyright convention by acquiring permission to showcase essential images for this nonprofit open-access editorial. However not all parties responded when approached or their contact details remained elusive. Images without permission have been drawn from the accurate representations of preserved or living biological structures, or those featured in brightfield/electron micrographs and photographs. All were in the public domain as accurate depictions of living or deceased structures or were drawn in conventional ways devoid of artistic expression or license. Such illustrations may be cited from multiple references, whereas permissions are attributed to the owner of copyright. The author and Reef Ranch Publishing Ltd have taken every precaution to comply with international copyright conventions.
To learn more, visit the ReefRanch website HERE
![]()

0 Comments