

By Joe Rowlett

The Angelfishes of the Pomacanthidae are undoubtedly some of the most universally popular species in the marine aquarium world. At the same time, their large size and showy patterns have made them some of the most well-documented and photographed of coral reef fishes… making it all the more surprising that so many grand mysteries yet remain concerning their evolutionary biology. Even relatively basic questions, like how many species there are and the geographic extent of their ranges, are shrouded in uncertainty. Chaetodontoplus is a moderately diverse genus of a dozen or so distinct phenotypes, several of which have had (and continue to have) a confused taxonomic history. By and large, these are fairly uncommon fishes found in moderately deep waters, typically below 20 meters, and often in siltier habitats than the coral reefs favored by most other pomacanthids. Uniquely among the angelfishes, this genus has a restricted distribution centered entirely on the West Pacific, absent even from the Melanesian waters of Vanuatu and the Fijian Plate. But, in spite of this provincialism, the group has managed to diversify into a bewildering amount of variation.

C. conspicillatus. Photo boris_44.
To date, no phylogenetic review or revision has ever been attempted for Chaetodontoplus. The group has been known since 1844, when C. septentrionalis was first described (within Holacanthus). The eminent Dutch ichthyologist Pieter Bleeker recognized the distinctiveness of this fish, and, in 1876, he established its current generic classification. The most prominent morphological trait for the genus is its unusually high number of lateral line scales (≈80, versus ≈40 in most other groups), with this multitude of minute scales lending a velvety appearance to these fishes. There doesn’t seem to be any common name applied to Chaetodontoplus, so I would like to suggest this feature be used to repurpose the term “Velvet Angelfishes” for the genus as a whole.
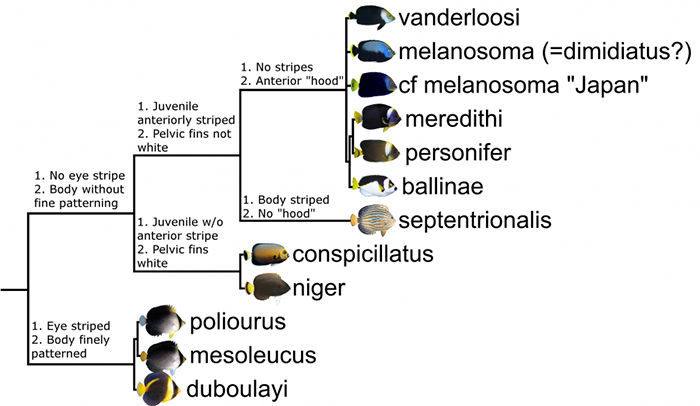
Phylogeny of Chaetodontoplus, inferred from morphology and cytochrome oxidase 1 (CO1).
The phylogenetic tree proposed here is based on a mixture of morphological and genetic study, as well as whatever piscine intuition I may have. Three main lineages become apparent: 1) Those species with a band running through the eye and with a heavily striated body. 2) Those which lack these features and whose juveniles lack a yellow stripe. 3) And a similar group whose members have a strong yellow stripe in the juveniles. These last two lineages are likely much more closely related to each other than either is to the first.
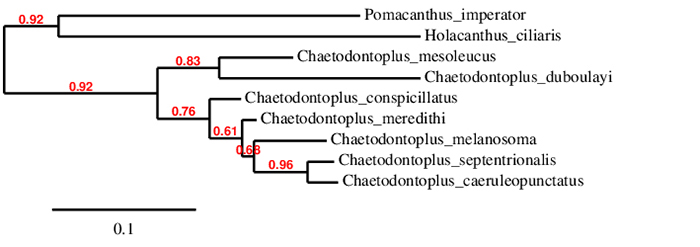
ML tree from available cytochrome b mitochondrial DNA.
Vermiculated, Greytail & Scribbled Velvet Angelfishes: C. mesoleucus, poliourus & duboulayi
Distribution map of C. mesoleucus, poliourus and duboulayi.
These three taxa are easily recognized as forming a distinct clade within Chaetodontoplus diagnosed by the prominent band running vertically through the eye, as well as the fine striations covering the sides of the body. Additionally, the juvenile coloration is more variably and diffusely patterned than is seen in most of its congeners.While many reef fishes stick to a consistent biogeographical endemism in the West Pacific, these three taxa follow the beat of their own ichthyological drummer. C. mesoleucus and poliourus range throughout the Coral Triangle, with the former found throughout the Indo-Philippines and the latter centered on the island of New Guinea. But poliourus also ranges further west along the Banda Arc, resulting in a wide area of overlap. This broad sympatry appears to maintain itself with assortative mating—that is to say, these two “species” are yet unknown to form mixed pairs. Genetic evidence has suggested upwards of 6% divergence between them (roughly treble the low-range threshold for recognizing distinct species), meaning these nearly identical fishes are split by a disproportionately deep genetic divide.

C. mesoleucus (Cebu, Philippines) and poliourus (Guadalcanal, Solomon Islands). Note the difference in caudal fin coloration. Photos by David Rolla & Mark Rosenstein.
Of particular interest is how poliourus appears to have expanded out from the typical New Guinean ecoregion seen in so many other fishes. Such Melanesian populations typically reach their westernmost extent in the waters around Northern Sulawesi and Halmaera, but in poliourus we find a population which has broadened to include Palau in the north and the entirety of the Banda Arc in the south. It’s not hard to image a fish with a relatively long pelagic larval stage journeying afar with help from the prevailing currents, and the flow in this portion of the Coral Triangle does match perfectly with what we see here—the Halmahera eddy driving fish northwards while currents in the Moluccas direct things south towards Bali. What remains to be answered, for this fish and for many others, is what barrier encouraged this speciation in the first place. What was the configuration (e.g. geography, ocean currents, habitat distribution) of the ancient West Pacific which fostered such distinct and isolated populations in Melanesia and the Indo-Philippines?

Male & female duboulayi. The striations become more linear in males, sometimes blending together (as in this large specimen). Photos by Michael K. Oliver & Pacific Island Aquapics.
Finally, C. duboulayi represents a close southerly relative of the previous species pair, and, despite some obvious differences in coloration, as well as it’s noticeably larger size, there is little doubt that this Aussie angel is a close relative of its more subdued congeners. In addition to the precision of their biogeographic fit and morphological similarities, genetic study (cyt b) supports the two together.

A tiny duboulayi. Note the similar, but wider, pectoral stripe relative to mesoleucus+poliourus. Photo by Petballoon.
This presents another biogeographic mystery, as duboulayi has an unusually extensive range which encompasses all the tropical coastlines of Australia. This raises the obvious question of how is it that C. duboulayi is able to seemingly maintain itself as a single cohesive “species” across the ecological barrier of the Torres Strait when so many other groups here have become isolated to the point of speciation? In fact, there is precisely such an example within this very same genus—C. meredithi and personifer. So what aspect of duboulayi’s biology or ecology might explain its continuing gene flow across Australia? Or might this actually be a case of a pair of cryptic species which have yet to be recognized? Extensive genetic study will be required to unravel the taxonomic mysteries of this group, but it’s worth noting that the remaining species of Chaetodontoplus all stick to the more familiar pattern of endemism seen in other coral reef fishes. There is a unique story to be told with these three species when it comes to the bigger picture of diversification and speciation in Indo-Pacific reef fishes. Despite these oftentimes being common and inexpensive fish in the aquarium hobby, they are nonetheless fascinating when viewed in their evolutionary context.
Conspicuous and Black Velvet Angelfishes: C. conspicillatus and niger
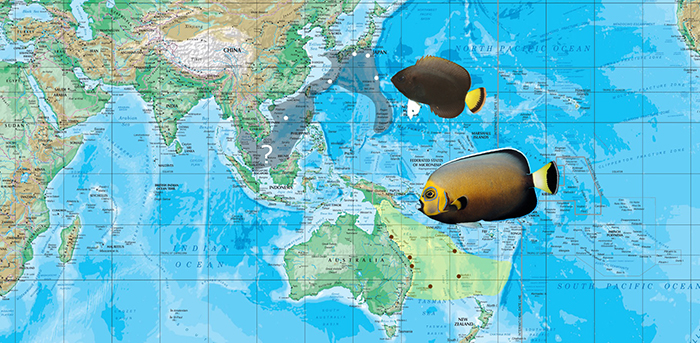
Distribution map of C. conspicillatus and niger.
In this small clade we find some of the rarest of the rare when it comes to aquarium fishes. The low abundance and restricted, subtropical range of C. conspicillatus has made it one of the most expensive species to be regularly exported, with an average retail price usually exceeding $2000. This is a trophy species for many aquarists, though, in this author’s opinion, one of rather depauperate aesthetic beauty. But the limited supply of “conspics” pales in comparison to it’s erstwhile sister species, C. niger, a pomacanthid of nearly unparalleled paucity.

A juvenile C. niger, seen at 28m in Sumisake, Japan. Photo by Suzuki Takahiro.
It remains entirely uncollected for the aquarium market and is known from only a handful of in situ observations. This fish appears to occupy a large range encompassing the waters of Japan and potentially all of the Southeast Asian mainland. One might understandably wonder how it can continue to be so rarely observed and so poorly documented given the extensive aquarium collecting and recreational diving conducted in this region. This seems to be due to a couple of factors: 1) C. niger favors deeper (50+ meter) reefs. 2) It occurs in silty, coral-poor habitats, away from where most collecting and diving is done.
C. niger has been photographed on rare occasions in shallower waters along the Japanese mainland. Only a couple of confirmed sightings are documented photographically, and these are of juveniles not more than a few centimeters in length. Adults are described to reach a much larger size (25cm) and to lose the black tip of the caudal fin. It has also been reported that they can take on more of a dark blue coloration, relative to the black of juveniles alluded to in the scientific name. Adults of C. niger are truly one of the last great angelfish discoveries yet to be made, right up there with rediscovering Apolemichthys guezei.

Juvenile C. conspicillatus lack the yellow pectoral band of its congeners. Photo by kaisuigyo-zukan.

A meredithi X conspicillatus hybrid, illustrating the genetic closeness of these taxa. Photo by Lemon Tea Yi Kai.
A couple traits of C. niger seem to align it with C. conspicillatus. Both possess white pelvic fins (though they are dark in juvenile conspicillatus), both lack any real patterning to their juveniles, showing only a diffuse dorsal lightening of the base coloration, and both have a black tip to the caudal fin, though this is apparently lost in adult niger. I find the juvenile commonalities to be particularly striking and supportive of these being a distinct pair occupying allopatric ranges. It’s interesting to note the great expanse of unoccupied habitat present in the Coral Triangle, suggestive of an antiequatorial distribution. Alternatively, this might also argue for these being more distantly related than I have indicated—two separate and distinct clades situated around Japan and Southern Australia respectively. A complete mitochondrial genome has already been sequenced for C. conspicillatus, so all that is required now is for C. niger to rear its melanistic head and let us grab a sample. If you happen upon this paradoxical piscine, be so kind as to grab a fin clipping.
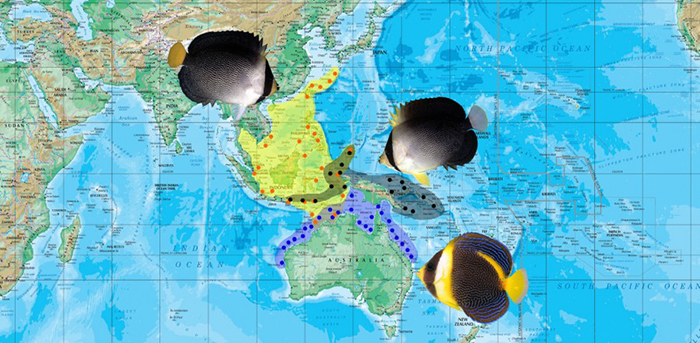
Half-black, Japanese, Vanderloos’, Meredith’s, Personifer & Ballina Velvet Angelfishes: C. melanosoma, cf melanosoma, vanderloosi, meredithi, personifer & ballinae

Distribution map of C. melanosoma, cf melanosoma, vanderloosi, meredithi, personifer and ballinae.
This diverse group has a confused and potentially incomplete taxonomy at the present time. It presents the typical regional endemism we see in many other Western Pacific reef fish groups, with the Japanese population appearing to be neglected and confused. What all of these taxa share is a similar coloration in the adults, with a predominantly black body and “hooded” head. Furthermore, there is often some amount of contrasting reticulation overlaid onto the face, which (in some hybrids) extends dramatically to cover much of the body. The juveniles are also informative, having a consistent pattern of a black base coloration and a distinctive yellow band running through the pectoral fin base. Note that this feature is present in their close relative, C. septentrionalis, but not elsewhere.

A) the typical “half-black” body and black caudal fin of the true C. melanosoma, from Anilao. B) the melanistic body of C. vanderloosi, from Eastern PNG. Photos by Oceanblue.
Differences between neighboring phenotypes are often quite subtle, though occasionally more abrupt. The Coral Triangle appears to have a pair of species, with melanosoma occupying most of the Indo-Philippines and vanderloosi known from the eastern tip of New Guinea. Despite the scarcity of recorded specimens, it can be assumed that this latter population will be found to occur throughout the Bismarck Archipelago and the Solomon Islands, though it’s surprising it has yet to be encountered given the thorough surveys which have been conducted in these regions.

The “dimidiatus” phenotype, from Raja Ampats and Palau Kawe (Bali). Photos by Gerry Allen & David R.
Things get more complicated in the west, where melanosoma is often split into a second species, C. dimidiatus. This latter taxon is said to be recognizable by its entirely yellow caudal fin (mostly black in melanosoma) and is documented from a limited region centered on West Papua, Halmahera and the Moluccas. It’s a confusing situation to explain, as there are no other large, pelagic-developing coral reef fishes which are endemic to just this region. Genetic studies conducted on clownfishes from this region have suggested a deep divide between the western and eastern populations of New Guinea, which might argue for the dimidiatus phenotype representing an unusual Western Melanesian species, or perhaps even a hybrid.Somewhat surprisingly, there aren’t any records of the typical black-tailed melanosoma phenotype having been documented from this region, which suggests that the yellow tail of dimidiatus might just be a localized variation. It’s important to note that juvenile melanosoma start off with only a submarginal black band in the caudal fin, the anterior portions later turning black. But specimens illustrating only partial darkening do exist, indicating that this is a more variable feature than has been appreciated. Note also the dimidiatus-like tail of Japanese specimens. Recall that poliourus+mesoleucus are both seen together in precisely this same region of the Coral Triangle, which is exactly what we would expect for two distinct species with overlapping biogeographies. Seeing as a similar situation doesn’t occur with melanosoma+dimidiatus, it seems implausible that the latter would be either a distinct “species” or even a hybrid, but this won’t be resolved without considerable genetic work.

C. cf melanosoma from Izu, Japan. Photos by ダイブログのブログ and kiss2sea.
That there might be an undescribed species of angelfish in Japan is almost ludicrous to think, but it seems that a confluence of confused taxonomy and extreme phenotypic plasticity has combined to leave a distinct species unrecognized here. Specimens collected from Cagayan, Philippines (likely representing the southernmost limit of this population) have been floating about the aquarium trade in recent years, occasionally being misidentified as the geographically distant C. vanderloosi. The two are in fact quite similar, with this Japanese fish differing most notably in having a predominantly yellow caudal fin. This is, of course, the exact feature I discussed as being untrustworthy as an identifying trait formelanosoma/dimidiatus, but the distinction of the Japanese faux-vanderloosi is supported by the wide geographic separation with respect to New Guinea. Both, in turn, can be identified from melanosoma and dimidiatus by their heavily black bodies.

Juvenile cf melanosoma. The partially yellow pelvic fins diagnose this fish from C. melanosoma, which has entirely black pelvic fins. Photos by kiss2sea and T. Takemura.
[Note that niger & melanosoma both translate etymologically as “black”. While the name “Black Velvet Angelfish” has long been applied to melanosoma, it is more fitting to the all-black body of niger, especially considering there are blacker species within the melanosoma complex].

Compare the variation in the facial markings of these two meredithi. Photos by Aquarise and Dave Harasti.
Further south, we find a pair of species occupying the traditional disjointed ranges along the tropical Australian coastlines. Originally considered a single species, the eastern phenotype was subsequently split (as C. meredithi) from the now Western Australian C. personifer. Because of this, the aquarium industry has perpetuated the common mislabeling of meredithia for the much more rarely-encountered personifer, or (slightly less egregiously) as the “False Personifer Angelfish”.
The two are distinctive at their extremes, but with a fair bit of confusing ambiguity present in many specimens. A good card-carrying meredithi should have a solid yellow tail and a blue patterning on the head. C. personifer, on the other hand, has a black bar in its caudal fin and a darkening to the face. It would be all well and good if these two phenotypes could stick to this arrangement, but, alas, it is not always so. Specimens of meredithi with hints of a caudal band or with a darkening of the head are known, just as there are personifer with mostly yellow tails.

Note the black face and black-banded caudal fin in C. personifer. Photo by Aquarise.
Which brings me back to my previous point concerning C. duboulayi, another taxon with a wide Australian range. What logic is there in treating duboulayi as a single species and splitting personifer/meredithi? Yes, there are more observable differences in the latter, but have we perhaps missed subtler differences amongst the western and eastern populations of the Scribbled Angelfish? The barrier which creates the common divide between the Australian coastlines is the repetitive appearance and disappearance of the Torres Strait through geological history. So, unless these two pan-Australian clades are drastically different in age, they should be expected to harbor similar amounts of genetic distance on either side of this barrier and should be given similarly distinct taxonomies.

The black caudal fin on this meredithi raises some questions… Photo by unknown.
Lastly, we come to the white whale of the velvet angelfishes—the enigmatic and alluring C. ballinae. It’s aberrant coloration can easily leave one confused as to its true affinities, but focusing on the similarities, rather than the differences, more clearly elucidates its evolutionary heritage. What we are seeing in this fish is almost certainly the result of its extreme endemism and low population numbers—a classic example of a “genetic bottleneck” creating unique traits.

C. ballinae is a sexy fish. Photo by unknown.
Originally discovered near Ballina, New South Wales in a fish trap placed in moderately deep waters, it has subsequently been discovered to be relatively common around Lord Howe Island and Ball’s Pyramid. It may yet be found in other nearby reefs, most of which are poorly explored at the depths one would expect to find C. ballinae at. The shallower waters of the Norfolk and Kermadec Ridges would be a good place to start looking.
The Ballina Angelfish is about as “book fish” as it gets, with only a handful of photographs in existence. Even Kuiter’s book on fishes from this region had to resort to a drawn illustration. Australia is particularly stingy in granting access to Lord Howe, so it may yet be a while before specimens can be collected and studied. Given the success had in captive breeding others in its genus, perhaps someday there can be captive-bred C. ballinae in the aquarium industry, as this may be the only fish capable of rivaling the leucistic beauty of Genicanthus personatus.
Blueline Angelfish (and it’s many hybrids): C. septentrionalis
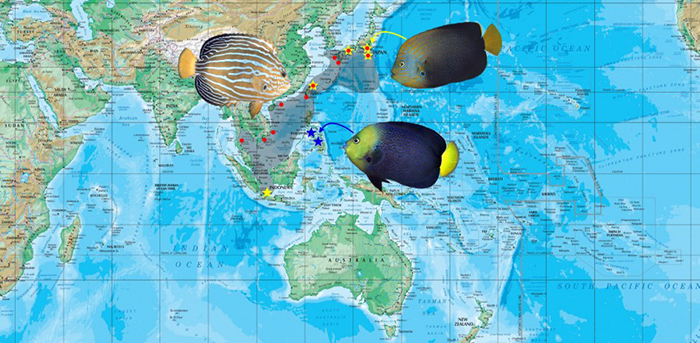
Distribution map of C. septentrionalis and hybrids.
There may be no greater confusion in all of reef fish taxonomy than that which pervades the many unusual phenotypes associated with C. septentrionalis. In its purest form, the Blueline Angelfish is an instantly recognizable species, completely unique within Chaetodontoplus for its prominent longitudinal lines. But, alas, septentrionalis is not content to breed amongst its own beautiful stock, and so it seeks the exotic allure of tastes forbidden. Photographic evidence has documented a fairly common pairing with the undescribed cf melanosoma in Japanese waters, which seems to produce a diverse array of hybrids.

Japanese & Vietnamese septentrionalis, showing striking variability. Photos by unknown.
A pair of names have gained some traction amongst hobbyist and scientific literature alike. C. chrysocephalus is a taxonomy given to those specimens displaying reticulations through the anterior portions of the body, far beyond what we typically would see in cf melanosoma. But, when we compare this to the greater amount of reticulation found in many septentrionalis specimens, it’s not hard to see how their hybrids might arrive at this appearance. A second purported species, C. cephalareticulatus, has also been described, though it’s not apparent how it is meant to differ from “chrysocephalus”, and most authors have tended to treat it as a synonym.

C. septentrionalis X cf melanosoma, in all its many forms. Photos by Nobukaza Koriya, Hiroshi Ueno, Tsutomu Asano, izuzaki, itadc.
When these various species were described, there was virtually no photographic documentation to inform their author’s taxonomic decisions, and a similar lack of preserved specimens did them no favours. What wasn’t apparent at the time was just how variable and ambiguous many of these specimens can appear. A vast continuum of phenotypes seems to exist, from those which could easily pass for the Japanese C. cf melanosoma, to highly-scrawled caerulean beauties that bear obvious resemblance to septentrionalis. What we are seeing can only be explained by a series of hybridizations and backcrossings between these two closely related taxa: C. septentrionalis and cf melanosoma. With the known existence of numerous in situ hybrid pairs, I think we can firmly put this confusing mystery to rest.

Some mixed “species” pairs, all from Japan. Photos by lodz, ダイブログのブログ, T. Ueno.
There is yet one more perplexing piscine to be found just to the south in the Central Philippines. The Bluespotted Angelfish (C. caeruleopunctatus) is an occasional aquarium export which goes for relatively modest sums of money. It’s affordability is seemingly out of step with how seldomly encountered and poorly known it is within the scientific community. The very peculiar biogeography of this fish should raise some alarms for those of you that have been paying close attention. There are species of coral reef fish endemic to the Philippines, but these are by and large known from groups with short or non-existent pelagic larval stages—there are otherwise no pelagic-developing species which exclusively reside here. So, how then do we explain the anomalous range of this bespeckled pomacanthid?

C. caeruleopunctatus (above) & “chrysocephalus”, strikingly similar. Photos by brucelee and kiss2sea.
Comparing the fine blue spots of the caeruleopunctatus phenotype to those seen amongst some of the “chrysocephalus” hybrids, we see a remarkable congruence. And when we observe how the range of septentrionalis is expected to reach into the Northern Philippines, a clear explanation begins to present itself. The Blueline Angelfish seems to envelop the waters around the Philippines (at least, it is expected to be found in these waters), and it would be no great shock for the occasional pelagic larvae to stray further afield than usual. Such a waif would have no recourse but to reproduce with its nearest sympatric relative (in this case, the true C. melanosoma). And this provides a plausible explanation for the “caeruleopunctatus” phenotype being nothing more than a hybrid, which differs from similar Japanese hybrids due to having a slightly different parentage.

Note the septentrionalis-like yellow caudal fin and the melanosoma-like coloration of the body in this juvenile “caeruleopunctatus”. Photo by Vessel.
To confirm this definitively, genetic sequencing (and lots of it) is required. But take a close look at the differences and similarities amongst these various fishes, and then compare that to their known ranges and the patterns of speciation we commonly see in coral reef fishes. There can be little doubt that the C. septentrionalis is an inordinately promiscuous little fishy.
Acknowledgments

More ballinae! Photos by volvox.
My thanks to Luiz Rocha and LemonTYK for their insightful commentary. Photo credits used in the maps are as follows: ballinae=Frank Baensch, “caeruleopunctatus”=brucelee, cf melanosoma=unknown, vanderloosi=unknown, “chrysocephalus”=kiss2sea,conspicillatus=boris_44, dimidiatus=Gerry Allen, duboulayi=Pacific Island Aquapics,melanosoma=oceanblue, meredithi=unknown, mesoleucus=John Ranall, niger=Suzuki Takahiro, personifer=Aquarise, poliourus=John Randall, septentrionalis=John Randall. Photo credit for C. conspicillatus=boris_44
References
Allen, G. R. and M. V. Erdmann. 2012. Reef fishes of the East Indies. Volumes I-III. Tropical Reef Research, Perth Australia. v. II: 425-855.
Allen, G. R. and R. C. Steene. 2004. Chaetodontoplus vanderloosi, a new species of angelfish (Pomacanthidae) from Papua New Guinea. aqua, Journal of Ichthyology and Aquatic Biology v. 8 (no. 1): 23-30.
Chan, W. L., 1969. Two new pomacanthid angelfishes from the Macclesfield Bank, South China Sea. Annals and Magazine of Natural History, ser. 13, 8: 325–334.
Kuiter, R. H. 1990. A new species of angelfish (Pomacanthidae), Chaetodontoplus meredithi from eastern Australia. Revue française d’Aquariologie Herpétologie v. 16 (no. 4) [1989]: 113-116.
Kuiter, R. H. 1993. Coastal fishes of south-eastern Australia. University of Hawaii Press, Honolulu. i-xxxi + 1-437.
Pyle, R. L., 2003. A Systematic Treatment of the Reef-Fish Family Pomacanthidae (Pisces: Perciformes). PhD thesis, University of Hawaii, Honolulu. xvi + 419 pp.
Randall, J. E. and L. A. Rocha. 2009. Chaetodontoplus poliourus, a new angelfish (Perciformes: Pomacanthidae) from the tropical western Pacific. Raffles Bulletin of Zoology v. 57 (no. 2): 511-520.
Yasuda, F. and Y. Tominaga. 1976. A new pomacanthid fish, Chaetodontoplus caeruleopunctatus, from the Philippines. Japanese Journal of Ichthyology v. 23 (no. 3): 130-132.

Beautiful genus! I am fond of Scribbled angelfish, I just wished that they didn’t cost so much! BTW, aren’t they being captive bred by Bali Aquarich?