


Fig 1. Siderastrea siderea ravaged by stony coral tissue loss (SCTLD) and black band (BBD) diseases. Image courtesy of The Florida Fish and Wildlife Commission ©.
This coral ailment was so devastating to certain scleractinian morphotypes that it not only prompted a national government rescue initiative, but it also forced ecologists to violate one of the canonical golden rules of conservation. Prolonged Noah’s Ark-type preservation and captive propagation with a view to populating the wild could prove disastrous, inasmuch as it does not exert the evolutionary-catalyst known as natural selection which may weaken species endurance. However, most fears have been belayed since the discovery of epigenetic modification and the recognition of gene introgression and phenotypic plasticity.
The author considered it time to renew his comprehension of this enigmatic illness because previous scientific insight could not fill a side of A4, which germinated an all-embracing article summarising the scientific community’s understanding of SCTLD.

Fig 2. The growth of a porcelain disc-mounted propagule of Pseudodiploria clivosa at a wild in situ nursery over six months. Image courtesy of The Florida Fish and Wildlife Commission ©.
The most informative white literature is the open-access seminal review of Papke and allies from 2024.
A disease instigating lesions of lost tissue on species of Montipora was reported in Hawaii in 2012, from which the bacteria Vibrio owensii were consistently isolated but failed to cause the same symptoms in healthy specimens (Ushijima et al. 2012). Stony coral tissue loss disease (SCTLD) has been rife in the wider Caribbean since 2014, which initiates as single or multiple foci of dead tissue that radiate outward to denude the entire colony in a few weeks. SCTLD is exceedingly contagious which appears to be spread via the water column or coral-to-coral or sediment-to-coral contact. Its environmental persistence is extraordinary, insofar as once established, it remains a continual threat (Weinberg 2018) irrespective of seasonal variance (Aeby et al. 2019; Alvarez-Filip et al. 2019; Muller et al. 2020; Estrada-Saldívar et al. 2021), which is redolent of abiotic causation. The progression of an illness orchestrated by biotic agents is intimately linked to their metabolism and that of their host, which is proportional to the temperature of the immediate surroundings in poikilotherms. Furthermore, corals associated with anthropogenically-developed coastal regions are the worst affected. Although less statistically conspicuous amongst the survey’s outbreak predictors, incidences of SCTLD were significantly higher within older marine protected areas and windward reefs. Intriguingly, this illness was absent in numerous predisposed corals on Banco Chinchorro’s series of isolated cays, where nominal anthropogenic activity combined with the surge of northbound Yucatan Current through a deepwater channel, insulates these habitats from the mainland (Fig 5.; Alvarez-Filip et al. 2022). Nevertheless, when the datasets from these reefs were eliminated from the prediction analyses, all other factors save coastal development became insignificant, where all sites were wind-exposed and holiday destinations (Alvarez-Filip et al. 2022).
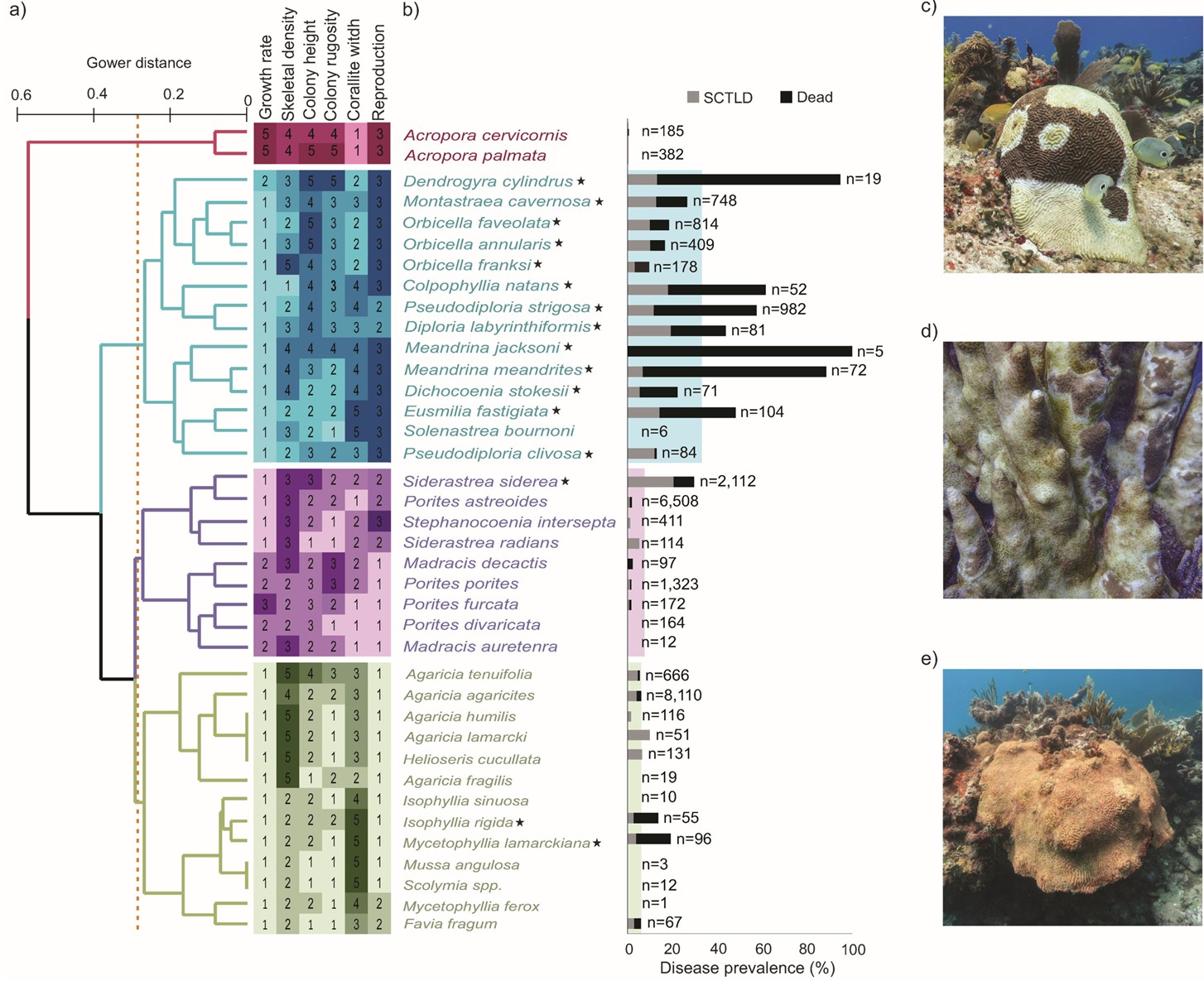
Fig 3. The species known to be vulnerable to SCTLD, their differences/similarities based on six aspects of their phenotypes and natural history, and the prevalence of SCTLD in each species in the Mexican Caribbean with reference to localized extinctions. (a) clustering dendrogram of the Gower distance analysis-based dissimilarity, not to be confused with a cladogram of ancestral lineage. The dotted line represents 65 percent dissimilarity, whilst the heatmap exemplifies the scoring of each trait which contextualizes the clusters that are not susceptible [red], those that suffer acute pathologies and swift mortality [blue], those that experience moderate disease [pink], and those that are barely affected [green]. (b) bar plots indicating the number of colonies of that species that are diseased or have succumbed. (c) Pseudodiploria strigosa with typical SCTLD; (d) Dendrogyra cylindrus most of which is algae-inhabited denuded skeleton, and (e) deceased Meandrina swathed in orange filamentous algae. [*] highly susceptible species, two of which, are in the barely affected category by virtue of their eradication and approaching localized extinction (Table 1.). Photography Lorenzo Alvarez . Analyses and data courtesy of Alvarez-Filip et al. 2022 and The Creative Commons Attribution License.
Before triggering an outbreak, SCTLD must be established in 0.05 to 0.1 percent of colonies in a locality (Dobbelaere et al. 2020; Papke et al. 2024) where four to 10 days incubation was required in recirculating systems (ex situ) before symptoms emerged (Aeby et al. 2019; Aeby et al. 2021; Meiling et al. 2021). However, computer modelling inferred incubation in the wild (in situ), can take anywhere from six days to six months (Dobbelaere et al. 2020; Guzman-Urieta & Jordan-Dahlgren 2021; Dobbelaere et al. 2022). Epidemiological analyses revealed clustering around infected colonies consistent with the spread of an infective live agent (Dobbelaere et al. 2020; Williams et al. 2021b; Truc et al. 2023). The findings of cluster analyses indicated the transmission of SCTLD is exceedingly inconsistent within and between regions and at different stages of an outbreak (Muller et al. 2020; Sharp et al. 2020; Aeby et al. 2021; Williams et al. 2021a).

Fig 4. Corallivorous foureye butterflyfish (Chaetodon capistratus) are recognised vectors of black band disease (BBD; Aebu & Santavy 2006) and are clearly partial to the tissue-boundaries of active SCTLD (Fig 3. c).
Dendrogyra cylindrus, Dichocoenia stokesii, Eusmilia fastigiata, and species of Meandrina appeared most susceptible. Mercifully, acroporids remain ostensibly unaffected (International Coral Reef Initiative 2019). Highly variable, the syndrome affected from less than 10 to 94 percent of the 21 susceptible Scleractinia. Over 50 percent of Meandrina meandrites of the family Meandrinadae and brain corals of the subfamily Faviinae succumbed over a few weeks, whereas pillar corals (Dendrogyra cylindrus) remain one of the most susceptible whose mortality has been documented at 80 percent (Alvarez-Filip et al. 2022).
The susceptibility tables 1 “a” and “b” feature species and their rates of tissue loss from various studies. Papke and allies remarked there is an urgent need for a global framework that standardises tissue loss analysis and SI units, as teams cannot compare and reproduce the results of percent tissue loss, unless datasets include the entire surface area of each colony. A direct measurement of the surface area lost over time would therefore be most appropriate, which would expedite the accrual of bioinformatics for sophisticated statistical analyses. Inter- and con-specifics, genotype, the conditions to which they are exposed, in situ versus ex situ appraisals, and the maturity of each colony impact susceptibility. Be that as it may, global paradigms must be established and preserved (Papke et al. 2024).
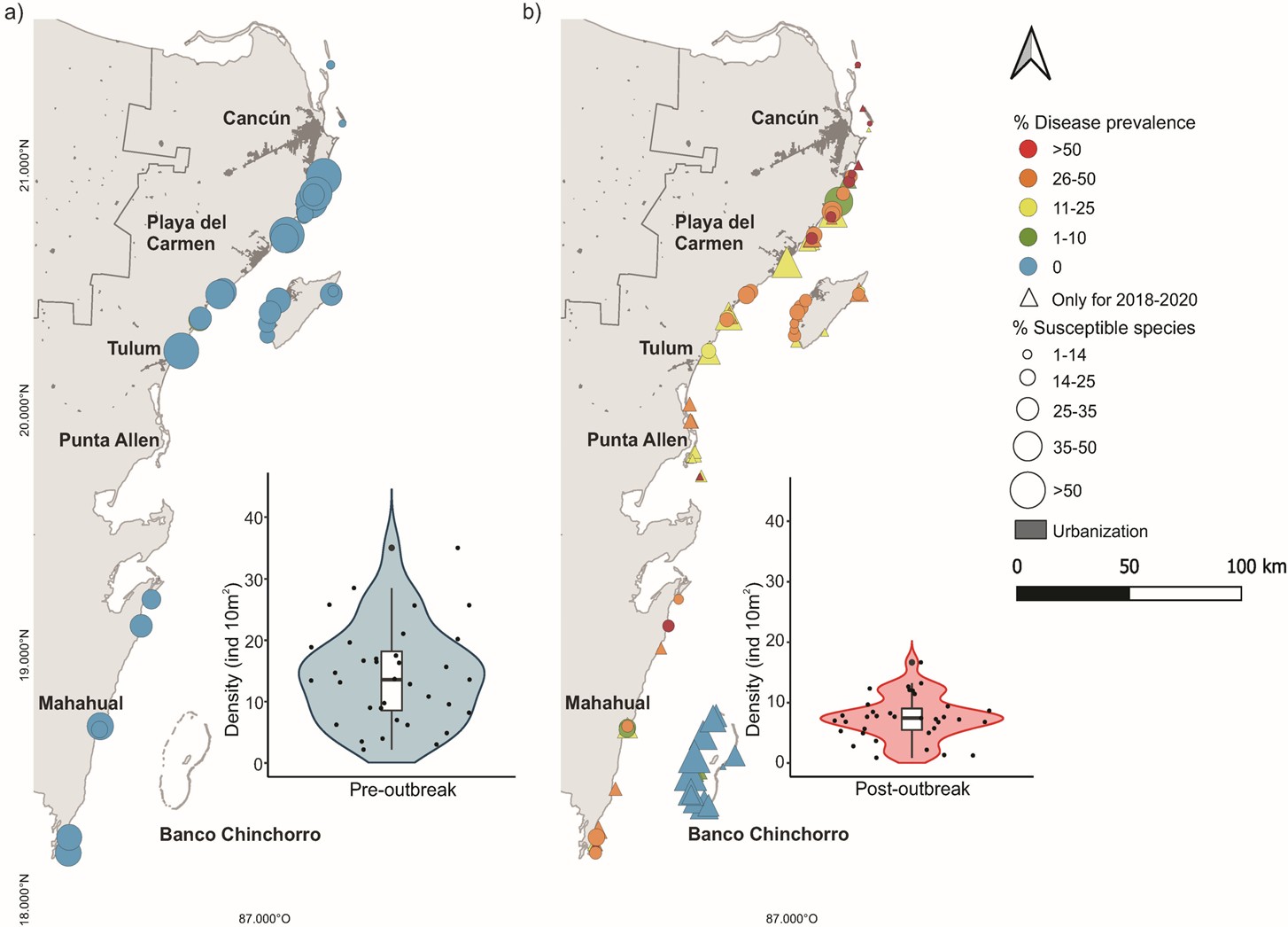
Fig 5. The prevalence of SCTLD in exceedingly susceptible species in the Mexican Caribbean. (a) a survey of white plague-like disease in species of coral that are extremely susceptible to SCTLD at 35 sites before the outbreaks of 2016 and 2017. (b) sites with populations of corals infected with SCTLD or those that have recently succumbed, in very susceptible species throughout, or following the outbreaks of 2018 and 2020. Circles represent the reefs that were surveyed before (a) and after (b) the emergence of SCTLD, whereas triangles mark the locations of the reefs that were surveyed during and after the outbreak. The size of each circle or triangle is proportional to the percentage of uninfected particularly-susceptible colonies at the site, out of all corals irrespective of species or susceptibility. (a) and (b) also feature the distributions of the densities of exceedingly vulnerable colonies across all sites for the pre- and post-outbreak periods respectively. Each point corresponds to a surveyed reef overlayed with box plots where the median is represented by the black horizontal, sandwiched between the first and third quartiles with 95th percentile whiskers. The shaded area of each violin plot illustrates the core density’s likelihood, whilst very susceptible species are those with more than 10 percent of infected colonies. Analyses and data courtesy of Alvarez-Filip et al. 2022 and The Creative Commons Attribution License.
Not only has the wider Caribbean been a notorious hotspot for emerging coral ailments (Green & Bruckner 2000; Harvell et al. 2007), but its reef community composition and dominant calcium carbonate depositors began to shift a decade or two before, from elkhorn (Acropora palmata; Appendix I), staghorn (A. cervicornis; Figs 22. & 26.; Aronson & Precht 2001; Alvarez-Filip et al. 2022) and their stable hybrid A. prolifera (Pinzón et al. 2010; Reich et al. 2021) to SCTLD-predisposed massive and boulder morphotypes. Hence the demographic of many of Florida’s reefs was reverting (Alvarez-Filip et al. 2022). Despite the wealth of keystone acroporan diversity responsible for most of the reef substratum accreted in the Indo-Pacific basin since the late Oligocene some twenty-eight to twenty-three million years ago (Wallace & Rosen 2006) upon which much of the local ecology relies (Zhao et al. 2021), staghorn and elkhorn and their hybrid, are the only Caribbean-wide congeners. Staghorn forms clumps or thickets of dispersed branches, whereas elkhorn erect sizeable, albeit dispersed, edifices but they have not recovered since they were listed as critically endangered in 2008 (Appendix I, photograph VI; Sutherland & Ritchie 2004; Hogarth 2006; Sutherland et al. 2011; Huang 2012). These taxa appear to devote, albeit not always (Toledo-Hernandez et al. 2023), few resources to constituent immunity (Brown et al. 2023) which may leave them vulnerable to pathogenicity and bleaching (Palmer 2018). They have thus suffered devastating losses (Goreau et al. 1998; Richardson et al. 1998; Ritchie & Smith 1998; Aronson & Precht 2001; Borneman 2002; Sutherland & Ritchie 2004; Hogarth 2006; Sutherland et al. 2011). Although staghorn is yet to be threatened to the brink, it too is comparatively rare, and as such, SCTLD has brought about a regional decline in calcite deposition by predisposing niches to inhabitation with encrusting corals that simply coat substratum or those with simple and low-reliefs. A predominance of Agaricidae and Porites astreoides could spell disaster in the face of predicted increases in sea level (Weil 2004; Perry & Alvarez-Filip 2019). See below: Other Considerations. According to historic and the current study’s surveys of reef community composition in Mexico’s Caribbean, there has been a 50.4 percent increase in these kinds of Scleractinia since 2016, which predates the first reports of SCTLD (Alvarez-Filip et al. 2022). However, it appears that SCTLD is contagious to these corals in recirculating systems. When Floridian P. astreoides shared water with the same ecosystem’s Montastraea cavernosa, Porites remained disease-free (Aeby et al. 2019); however, four years later, the disease was transmitted to this species from the U.S. Virgin Islands when it shared ex situ water with infected Diploria labyrinthiformis from the same region (Meiling et al. 2021). The infectious agent(s) which is suspected to be a bacterium or a consortium thereof (Aeby et al. 2019; Neely et al. 2020; Ushijima et al. 2020; Shilling et al. 2021; Walker et al. 2021; Forrester et al. 2022) and species of coral likely evolve to be geographically and regionally distinct (Veron 1995, cited in Arrigoni et al. 2016; Ramirez-Portilla et al. 2022). Exceedingly susceptible taxa experience accelerated rates of tissue loss and heavy mortalities, whilst those that the disease barely affects experience slow progression and chronic pathologies. Similarly, there are species of coral that suffer moderate-disease and -paced-pathogenesis. D. labyrinthiformis falls into the former category, whereas M. cavernosa could be classed in the latter. Hence these corals may have a respective greater or lesser propensity to spread the disease (Papke et al. 2024). No ex situ experimental challenges have been performed on corals of the family Agaricidae but a wealth of studies have documented their mild lesions in the wild (Brandt et al. 2021; Combs et al. 2021; Costa et al. 2021, cited in Papke et al. 2024; Heres et al. 2021; Williams et al. 2021a, cited in Papke et al. 2024; Hayes et al. 2022, cited in Papke et al. 2024). Hence despite the acute prognosis of figure 6. C, Agaricidae and Porites astreoides should be regarded as barely susceptible (Papke et al. 2024). Moreover, to date, no peer-reviewed literature or experimental datasets assert SCTLD does not infect species of Acropora (Papke et al. 2024).
As SCTLD spreads into a new territory, the most vulnerable species such as Dendrogyra cylindrus, Dichocoenia stokesii, Eusmilia fastigiata, and Meandrina meandrites typically suffer close too or 100 percent mortality within a matter of weeks. Then as the epizootic attains prevalence in the moderately susceptible, there is a concomitant deceleration until the worst affected of that class succumb whilst others barely subsist. At this juncture, the ailment slows to chronically affect the most resistant (Papke et al. 2024). Therefore, SCTLD is somewhat self-limiting inasmuch as once a fire’s fuel is all but spent, it smoulders. However, mass extinctions are anything but favourable prognoses. Like all populations and their interactions with pathogens, conspecific susceptibility varies (Aeby et al. 2019; Meiling et al. 2021) where long-term survival of the host and its pathogen is mutually beneficial.
Unsurprisingly, efforts to isolate a definitive SCTLD-causing microorganism(s) have found the ubiquitous microbiome affiliates bacteria, bacteriophages, and viruses, while the syndrome exerts physiological impacts on host and zooxanthellae (Aeby et al. 2019; Landsberg et al. 2020; Work et al. 2021). Most studies have been conducted in Mexico and Florida where SCTLD has extended its host range to nearly 30 species (Estrada-Saldívar et al. 2021, cited in Papke et al. 2024). Those affected include: Colpophyllia natans, Dendrogyra cylindrus, Diploria labyrinthiformis, Dichocoenia stokesii, Eusmilia fastigiata, Isophyllia rigida, Montastraea cavernosa, Meandrina meandrites, M. jacksoni, Mycetophyllia lamarckiana, Orbicella annularis, O. faveolata, O. franksi, Pseudodiploria clivosa, P. strigosa, and Siderastrea siderea (Fig 3.; Alvarez-Filip et al. 2022). P. strigosa was previously considered a robust species resistant to many of the stressors associated with climate change (De Bakker et al. 2016).
Although the biotic-instigator(s) remains enigmatic, there is an earnest desire to isolate the cause. Ex situ challenges in recirculating systems are redolent of the transmission of an infective organism(s; Dobbelaere et al. 2020; Williams et al. 2021b; Truc et al. 2023) in water and sediment (Muller et al. 2020; Dobbelaere et al. 2022; Studivan et al. 2022a; Studivan et al. 2022b) with multiple candidate vectors (Noonan & Childress 2020; Rosales et al. 2020; Studivan et al. 2022a; Studivan et al. 2022b).
It appears that the aetiological agent or agents, manipulate the coral to express genes implicated in pathogen recognition and those that reprogram innate immunity (Traylor-Knowles et al. 2021).
Our understanding of disease is evolving because many do not originate from the exploits of a primary pathogen, which do not merely interact with the host and its immune system, but with the multipartite holobiont and its immediate surroundings. Illnesses appear a consequence of a dynamic process whereby microeukaryotic and prokaryotic symbionts, commensals, and their viruses, initially attempt to outcompete and eradicate invading microorganisms by liberating antimicrobials and modifying their community compositions and metabolism. Cellular trauma that exceeds their restorative potential demands participation of adaptive and innate immunity. Host defence transforms the conditions within its microbiomes to curtail spread and purge disease-causing agents, which leads to transitory and intended states of microbial-purging known as dysbiosis. The pathobiont is the consequence of the host’s altered constituent physiology and its now hostile microniches (pathobiomes), and its modified community and metabolic function (pathobolome) which we hitherto called disease (Bass et al. 2019). Furthermore, prokaryotic communities play a key role in recouperation from bleaching (Peixoto et al. 2017; Santoro et al. 2021). Numerous studies have found that the microbial community compositions of coral holobionts shift in response to disease (Rosales et al. 2020; Becker et al. 2021; Clark et al. 2021; Thome et al. 2021; Work et al. 2021; Rosales et al. 2023), whose profiles may realign before the emergence of SCTLD (Huntley et al. 2022). Moreover, transcriptome modification occurred in the “algal” partners of exposed but non-diseased colonies (Beavers et al. 2023).
Even when Koch’s postulates or extensive genetic analyses have confirmed a particular pathogen or pathogens cause a specified disease, later research has found that strain(s) or species absent in lesions or present in healthy specimens (Porter et al. 2001; Cooney et al. 2002; Pantos et al. 2003; Sunagawa et al. 2009; Casamatta et al. 2012; Ho 2013; Soffer et al. 2013; Joyner et al. 2015; Sutherland et al. 2016). A prime example is Aspergillosis of Caribbean sea fans where the scientific community was convinced that the fungus Aspergillus sydowii caused the disease. The illness prompted a profound and extraordinary immune response that induced melanisation cascades and the overproduction of sclerites to the extent it caused discolouration and galling. One can see that this might be mistaken for a pathogen reprogramming the immune system. It was later postulated that a shift in the community composition of the coral’s mucosal microbiome was largely responsible for disease (Toledo-Hernández et al. 2008; Tait et al. 2010; Cardenas et al. 2011; Egan & Gardiner 2016; Morrow et al. 2018). Anomalous elevated temperatures induce the lytic cycle in viruses that infect corals and their zooxanthellae, which frequently opens niches for opportunistic microorganisms to inhabit and cause pathology (Lesser et al. 2007; Hall et al. 2018; Morrow et al. 2018). And as already mentioned, rates of microbial and coral ana- and meta-bolism are typically positively correlated with temperature. Exceptions occur when a nonpermissive degree is reached at which an organism can no longer survive, or when immune resources are sequestered to dissipate stress from surplus irradiance and heat that might otherwise be used for growth. Hence Acropora cervicornis grow more rapidly in winter (Toledo-Hernandez et al. 2023). Remarkably, the severity and virulence of SCTLD is not influenced by temperature but what appears to be host susceptibility which is more indicative of environmental causation. However, using an inoculum to successfully recreate the same disease in healthy specimens is not (Muller et al. 2020; Dobbelaere et al. 2022; Studivan et al. 2022a; Studivan et al. 2022b). All that is required in this instance to satisfy Koch’s postulates is the consistent isolation of a specific candidate pathogen from all lesions and the inoculum, and the artificial uncontaminated culture of that organism, which science has yet to accomplish, and as such, all we know is that the symptoms appear the same, but that does not substantiate we are dealing with the same illness (Appendix I, photograph V; Papke et al. 2024). Tissue can die and slough from corals for all kinds of reasons least of all, an intrinsic cellular response called preprogrammed cell death which is synonymous with apoptosis (Godar 1996; Ainsworth et al. 2007). Between 21 and 56 percent of wild scleractinian tissues are in a continual state of regeneration (Palmer et al. 2011; Palmer et al. 2012; van de Water et al. 2015).

Fig 6. Photography of Caribbean corals exhibiting putative lesions of SCTLD: [A] Dendrogyra cylindrus; [B] Siderastrea siderea; [C] Agaricia agaricites; [D] Pseudodiploria strigosa; [E] Colpophyllia natans; [F] Diploria labyrinthiformis; [G] Eusmilia fastigiata; [H] Dichocoenia stokesii, and [I] Meandrina meandrites. Photograph D, Dan Mele, the remainder, Sonora Meiling. Courtesy of Papke et al. 2024 and The Creative Commons Attribution License.

Fig 7. The gross morphology of lesions tentatively identified as SCTLD where less than 1 cm recently-denuded bands indicate typically non-lethal chronic pathologies [A & E]; peculiar dark possibly melanised regions on Siderastrea siderea around the periphery of surviving tissue [A]; 1 to 5-cm banded examples of moderately-progressing subacute lesions that eventually lead to colony demise [B & F]; whereas over 5-cm bands suggest acute swift pathologies typically instigating mortality [C & G], whilst [D & H] are examples of bleached tissue immediately ahead of the advancing front. Photograph H, Rosmin Ennis, the remainder, Sonora Meiling. Courtesy of Papke et al. 2024 and The Creative Commons Attribution License.
Upon occasion, tissue loss has simply ceased (Aeby et al. 2021, cited in Papke et al. 2024) whilst opportunistic coinfections that hasten progression cannot be ruled out (Ushijima et al. 2020).
Other Considerations
Reefs can drown in regions where seawater is rising disproportionately, the seafloor is undergoing accelerated subsidence, the calcium carbonate substratum is eroded by rising atmospheric CO2 and oceanic acidification or endogenous hydrogen sulphide, or enriched nitrogen and phosphorus fuel the growth of solubilising microalgae (Perry et al. 2014; Molina-Hernández et al. 2020; Alvarez-Filip et al. 2022). Unless reef vertices are rapidly developed by skeletally-complex zooxanthellate Scleractinia, intense light-demanding hermatypes can find themselves plunged into inhospitable mesophotic depths. The euphotic zone where light-dependent organisms thrive, extends in clear water to merely 37 metres on The Great Barrier Reef (GBR; Michael et al. 2011) where Atoll formation is constrained insofar as the seafloor of northeastern Australia does not subside (Purdy & Winterer 2005).
Increased expanses of bare skeleton can recruit fleshy or filamentous algal turfs which curtail the settlement of coral planulae (Berger, personal communication; Bulleri et al. 2018; Schmitt et al. 2019) and liberate coral larva-killing allelopathic biochemicals (Birrell et al. 2008). Once these organisms are established, reef communities shift from coral dominated to less productive ecosystems inundated by macroalgae which retain an inertia and are unlikely to revert (De Bakker et al. 2016; Alvarez-Filip et al. 2022; Smith 2023). Moreover, such is the ecology of several Caribbean reefs since the localised extinction of prolific herbivorous long-spined black sea urchins (Diadema antillarum; Lessios et al. 1984; Lessios 2016; Munn 2019).
Figure 4. features a corallivorous foureye butterflyfish (Chaetodon capistratus) which are recognised vectors of black band disease (BBD; Aebu & Santavy 2006). Richardson and allies isolated a variant BBD consortium in the Florida Keys and Bahamas whose lesions were pervaded by the cyanotoxin microcystin (Richardson et al. 2007). The lethality of this hepatotoxin is enough to dissuade the most voracious of Chaetodon capistratus, yet the microbial mat of BBD did not generate microcystin elsewhere (Richardson et al. 2007). Clear evidence indicates that SCTLD is communicated through the water column (Weinberg 2018; Alvarez-Filip et al. 2022). Given that SCTLD can develop from multiple foci, intuition infers the feeding habits of these butterflyfish are somehow involved in the infective process (Noonan & Childress 2020). Yet equally, they may be bioremediatory as they nibble lesion boundaries (Fig 3. c) while they remain implicated in recuperation (Titus et al. 2022). Although these fish do appear to assemble around and preferentially target SCTLD-infected colonies in the wild (Noonan & Childress 2020), captive foureye butterflyfish do not demonstrate a statistically significant dietary preference for diseased colonies, whilst their corallivory does not increase the transmission of SCTLD (Titus et al. 2022).
Intriguingly, merely 241 colonies out of 29095 exhibited the clinical signs of other coral diseases during Alvarez-Filip and colleagues’ year-round survey of an 18 to 21o North 450-km tract of the Mexican Caribbean from July 2018 to January 2020 (Alvarez-Filip et al. 2022). Conspicuous by their absence in the months of summer and autumn where they are at their most virulent, perhaps future work can investigate why, at least in this instance, SCTLD appears to palliate or mask other coral disease. Could it be, SCTLD outcompetes other coral ailments to extinction.
25 out of 48 species affected by SCTLD in Mexico’s Caribbean, were Robusta mid-to-large massive morphotypes with dense skeletons that broadcast gametes (Alvarez-Filip et al. 2022).
The species that suffered the most devastating losses had similar rates of growth, skeletal density, colony height and unevenness of surface (rugosity), corallite width, and reproductive mode (Fig 3.). A loss of genetic heterogeneity that results from mass extinctions of subpopulations weaken species endurance and adaptability, which threatens survival in an everchanging environment. Be that as it may, numerous kinds of juvenile colonies appear unaffected, and adults with expanses of denuded skeleton are able to sexually reproduce (Alvarez-Filip et al. 2022). Their gametes can also cross fertilise those from undiseased colonies (Quiroz et al. 2023). Nevertheless, juvenile Colpophyllia natans or Diploria labyrinthiformis appear to lose tissue faster (Williamson et al. 2022) or slower (Meiling et al. 2021, cited in Papke et al. 2024) than their mature cohorts. Yet excluding rapid tissue necrosis (RTN; Bythell et al. 2004), SCTLD advances at an alarming rate compared with other diseases like white plague that cause tissue loss in corals of the wider Caribbean (Clemens & Brandt 2015; Papke et al. 2024).
The diversity-disease or dilution effect hypothesis (Keesing et al. 2006; Huang et al. 2016) proposes infections are less prevalent amongst more diverse populations of corals, whilst SCTLD appears comparatively less widespread where scleractinian diversity is constrained (Muller et al. 2020; Costa et al. 2021; Williams et al. 2021a). The prevalence of SCTLD in the year following its emergence typically ranges from 16 to 57 percent (Precht et al. 2016; Alvarez-Filip et al. 2019; Estrada-Saldıvar et al. 2020; Aeby et al. 2021, cited in Papke et al. 2024; Brandt et al. 2021; Estrada-Saldıvar et al. 2021; Williams et al. 2021a; Hayes et al. 2022) accompanied by a loss of up to 62 percent coral cover (Brandt et al. 2021; Estrada-Saldıvar et al. 2021; Heres et al. 2021; Kolodziej et al. 2021; Williams et al. 2021a; Hayes et al. 2022). Highly and moderately susceptible species that are rapidly eliminated tend to be a minority of reef communities (Costa et al. 2021; Neely et al. 2021a), and thus SCTLD has a pronounced effect on diversity (Estrada-Saldıvar et al. 2020; Brandt et al. 2021; Heres et al. 2021; Hayes et al. 2022). It does not, however, typically denude vast areas of a reef because of the comparatively low-footprint of the rare species it kills (Costa et al. 2021). Its impacts are frequently underestimated because tiny and juvenile colonies are typically excluded from surveys (Williamson et al. 2022). Near- and off-shore prevalences differ (Rippe et al. 2019, cited in Papke et al. 2024; Muller et al. 2020; Williams et al. 2021a). Older or larger colonies may be more vulnerable (Sharp et al. 2020; Williams et al. 2021a); hence where the illness has been rife for some time, merely small examples of highly susceptible species may be found (Aeby et al. 2021; Papke et al. 2024). Although several studies have reported that SCTLD remains unaffected by seasonal changes in water temperature (Aeby et al. 2019; Muller et al. 2020; Hayes et al. 2022; Truc et al. 2023, cited in Papke et al. 2024) others have documented symptoms subside during anomalous elevated temperatures (Meiling et al. 2020; Sharp et al. 2020; Williams et al. 2021a) which may be a consequence of reduced populations of zooxanthellae (Papke et al. 2024).
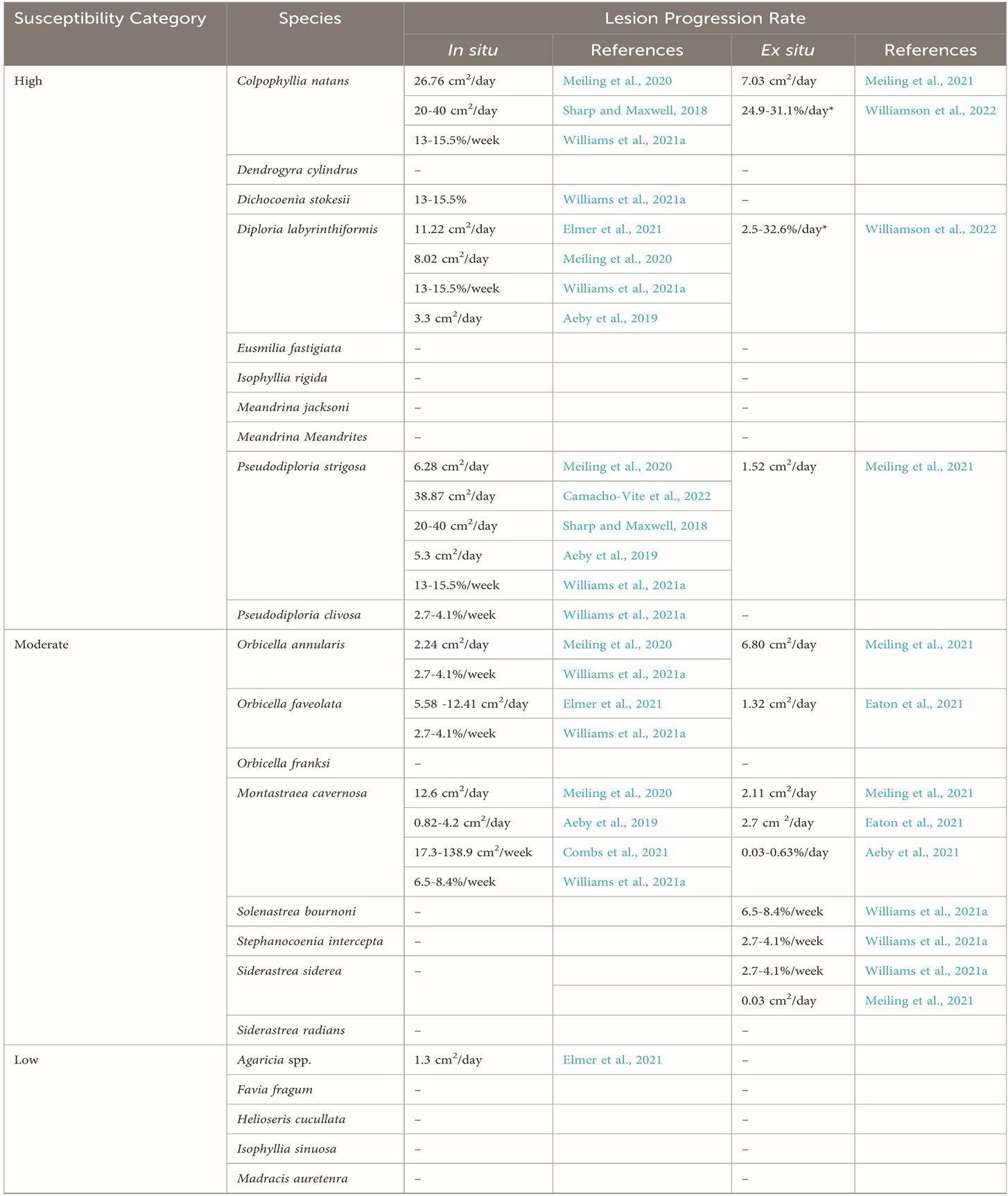
Table 1a. Categories of susceptibility to SCTLD and disease progression: [-] no published data; [*] appraised on tiny juveniles. There is an urgent need for a global standard for analysing and measuring tissue loss (Papke et al. 2024). Courtesy of Papke et al. 2024 and The Creative Commons Attribution License.
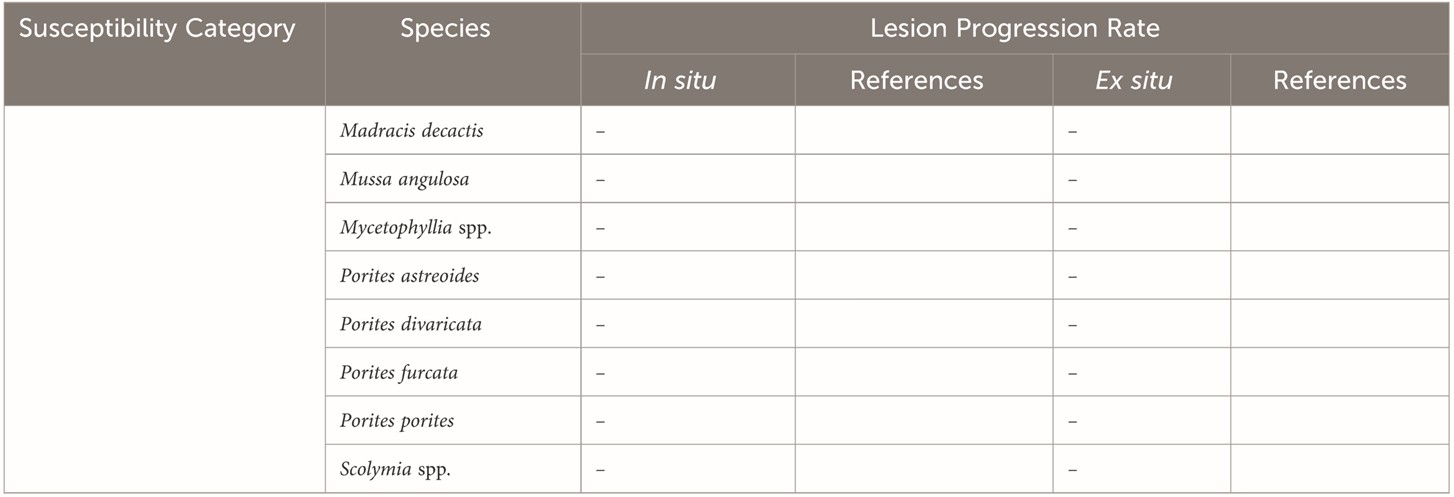
Table 1b. Continuation of low susceptibility to SCTLD: [-] no published data; [*] appraised on tiny juveniles. Courtesy of Papke et al. 2024 and The Creative Commons Attribution License.

Fig 8. A subacute to acute instance of SCTLD on Pseudodiploria strigosa in the U.S. Virgin Islands with a band approaching 5 cm of recently denuded skeleton. Courtesy of Dan Mele ©.
Pathogens do not intend to cause mortality insofar as death of their host results in their demise. Therefore host/parasite interactions eventually attain a form of equilibrium, so pathogenesis instigates merely moderate discomfort and malaise (Penczykowski et al. 2015). Newly evolved pathogens or those that have extended their range to infect a new kind of host, have yet to attain this kind of stability. SCTLD appears self-limiting in two ways: it rapidly eradicates its preferred hosts which lessens diversity which in turn curtails its prevalence. This could indicate the aetiological agent(s) has only recently been introduced into the marine environment, has just evolved, or corals are not its conventional host.
Coral Immunity and SCTLD
Very little is known about the interrelations of SCTLD and coral immunity (Papke et al. 2024), and although founded on the discoveries in peer reviewed white literature, much of our understanding of scleractinian defence is theoretical and comparatively young.
The immune systems of all organisms must distinguish self from nonself to minimise potentially-harmful autoimmunity (Kvennefors et al. 2008). However, this concept may be outdated insofar as microbiomes that comprise microeukaryotes and prokaryotes are integral components of the holobiont and its defense (Palmer 2018). Zooxanthellate Scleractinia (hermatypic corals) do not have cell-mediated (adaptive) immunity driven by ultra-precise molecular shape recognition, which detects foreign antigens through the unification of exacting complementary-shaped epitopes and paratopes found on antigens and antibodies. Yet corals retain a less fastidious arm of defence called innate immunity that relies upon comparatively imprecise pattern recognition receptors (PRRs) and fundamental clotting and biocidal competencies, which is both intracellular and extracellular and depends upon the transcription of a suite of immune-associated genes (Palmer et al. 2008; Palmer et al. 2010; Palmer et al. 2012; van de Water et al. 2015; Toledo-Hernandez et al. 2023). It was evident from studies that microorganisms in the mucosal microbiome played a vital role in colony defense, which supported the longstanding corals are essentially rudimentary paradigm, so evidence to the contrary was somewhat astounding.
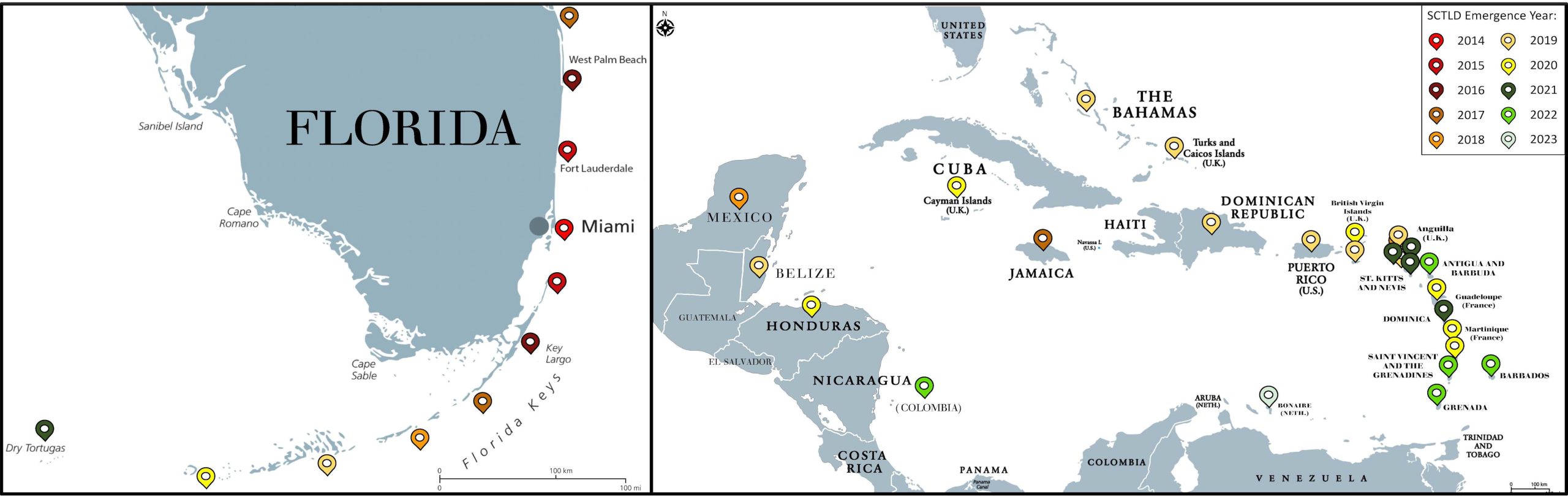
Fig 9. Maps of Florida and the wider Caribbean illustrating the year and site that SCTLD emerged. Courtesy of Papke et al. 2024 and The Creative Commons Attribution License.
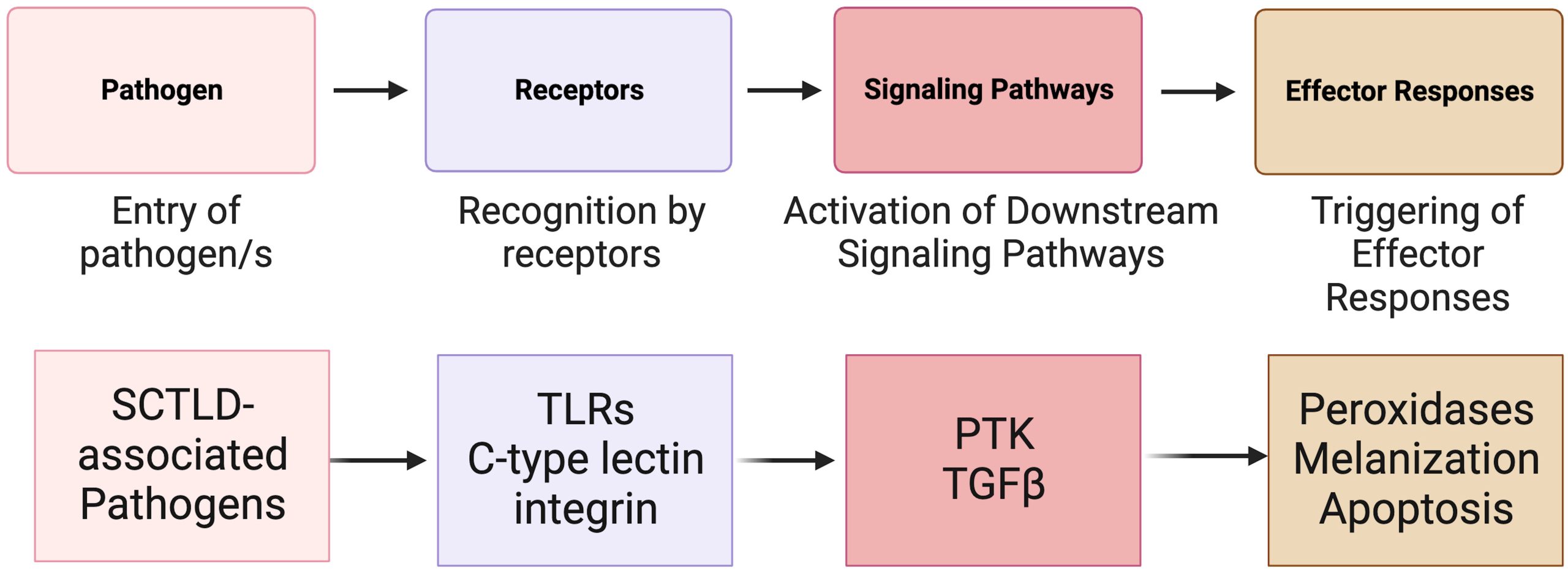
Fig 10. A fundamental illustration of the key elements of invertebrate immunity and the sequence of events involved with generalised contagion of an infective organism (top) and the corresponding SCTLD-triggered receptors, induced cytokines, and mechanisms used by host immunity. [TLRs] toll-like receptors; [PTK] the protein tyrosine kinase pathway, and [TGF] the transforming growth factor-beta pathway. Composed by Papke and allies from the information within Traylor-Knowles et al. 2021 and Beavers et al. 2023. Courtesy of Papke et al. 2024 and The Creative Commons Attribution License.
Representatives of 60 or more viral families have been isolated from corals (Ambalavanan et al. 2021) where bacteriophages are likely the most plentiful (Wegley et al. 2007; Nguyen-Kim et al. 2015). Lysogenic viruses are integral to the genomes of zooxanthellae which undergo lytic induction when nutrients are enriched or when temperature and/or irradiance are elevated, and thus lysogeny expedites bleaching (Fig 11.; Munn 2019). An upsurge of virus-like particles (VLPs) and insult to the integrity of zooxanthellae that ultimately leads to degradation, are commensurate with the onset of SCTLD (Work et al. 2021; Papke et al. 2024). These VLPs were tentatively classified in the family Alphaflexiviridae (Work et al. 2021) where genomic studies detected two of its members in healthy, suspected-exposed, and SCTLD-infected corals (Veglia et al. 2022). However, the abundantly-ubiquitous nature of marine viruses means their presence does not necessarily indicate they are involved in an infective pathogenic process (Howe-Kerr et al. 2023). Beavers and collaborators found that transcripts of Rab7 accompany the disintegration of the “algal” symbiome, the host’s photosynthate-subsidised mucosa undergoes an operational shift, zooxanthellae are damaged by viruses, and the phylogeny of the coral’s “algal” partners influenced the virulence of SCTLD (Beavers et al. 2023). However, the dynamic communities of the holobiont are critical components of coral immunity which donate often vital metabolic by-products. Their meta-genomic and -bolomic plasticity appears pivotal in the early contagion of infective organisms (Boilard et al. 2020) and are thus undoubtedly embroiled during colonisation of the microorganisms that “cause” SCTLD and the development of a pathobiome (Bass et al. 2019).
A plethora of studies have investigated how the holobionts of corals respond to SCTLD (Rosales et al. 2020; Becker et al. 2021; Clark et al. 2021; Thome et al. 2021; Work et al. 2021; Huntley et al. 2022; Rosales et al. 2023) where community shifts may be driven by members of the bacterial order, Flavobacteriales (Rosales et al. 2023). Diseased corals differentially express hundreds if not thousands of genes (Traylor-Knowles et al. 2021) many of which encode antioxidants (Jin et al. 2016). Attributes of the immune system include collagens, peroxidases, bax- and fibrinogen-like proteins, transforming growth factor beta (TGF-β), and tyrosine kinases (Traylor-Knowles et al. 2021), which assist intercellular communication and tissue maintenance through the phosphorylation of tyrosine residues using the terminal phosphate of adenosine triphosphate (ATP; Hubbard & Till 2000). Tyrosine kinases and their downstream cascades elicit immune responses in Orbicella faveolata (Traylor-Knowles et al. 2021), whereas peroxidases are upregulated in the same species and Montastraea cavernosa suffering from SCTLD which are plentiful in corals that have lost tissue (Halliwell & Gutteridge 2015; Abbas et al. 2019).
Fig 11. The lytic life cycle of T4-like bacteriophages.
TGF-β is a multifunctional signalling cytokine (Chaudhury & Howe 2009) whose expression appears central to early zooxanthellae recruitment and immunity, because TGF-β-disrupted mutant spats (accreted first polyps) or adults could not acquire “algal” partners or generate the cellular toxin (cytotoxin) nitric oxide (NO; Berthelier et al. 2017). Hence the inhibition or suppression of TGF-β appears a prerequisite for an efficacious defence (Fuess et al. 2020). The TGF-b pathway is suppressed before immunity responds to SCTLD where the expression of integral protein SMAD6 was significantly downregulated (Traylor-Knowles et al. 2021, cited in Papke et al. 2024; Beavers et al. 2023). Diminishment of SMAD6 is associated with increased incidences of apoptosis in human cells (Jeon et al. 2008). Signal transduction is compromised in holobionts with advanced SCTLD (Papke et al. 2024).
Coral receptors continuously monitor the homeostasis of the holobiont and respond to a- and -biotic disturbances using their downstream molecular cascades, effectors, biomolecular signals, and antimicrobial by-products, whilst recognition by the microbial network modifies cross-anatomy gene expression in order to stabilise associations and reinforce eubiosis (Palmer & Traylor-Knowles 2012; Preston 2015; Palmer 2018; Boilard et al. 2020; Santoro et al. 2021).
Coral mucus, tissues, and skeletons are inhabited by dynamic communities of immeasurable interdependent microorganisms which thrive on the metabolic by-products of their neighbours, where commensals and symbionts undertake vital housekeeping. Constituent immunity must optimise holobiont associations through winnowing and recruitment while tolerating healthy populations. Integrated stress responses (ISRs) exploit protective extracellular oxidative bursts of reactive oxygen species (ROS) which have until late become biomarkers of thermal stress (Nielsen et al. 2018; Palmer 2018). The danger model of immune activity proposes molecules associated with cellular rupture like heat-shock proteins, uric acid, and other reactive molecules alert immunity to impairment which devises an operational response (Palmer 2018).
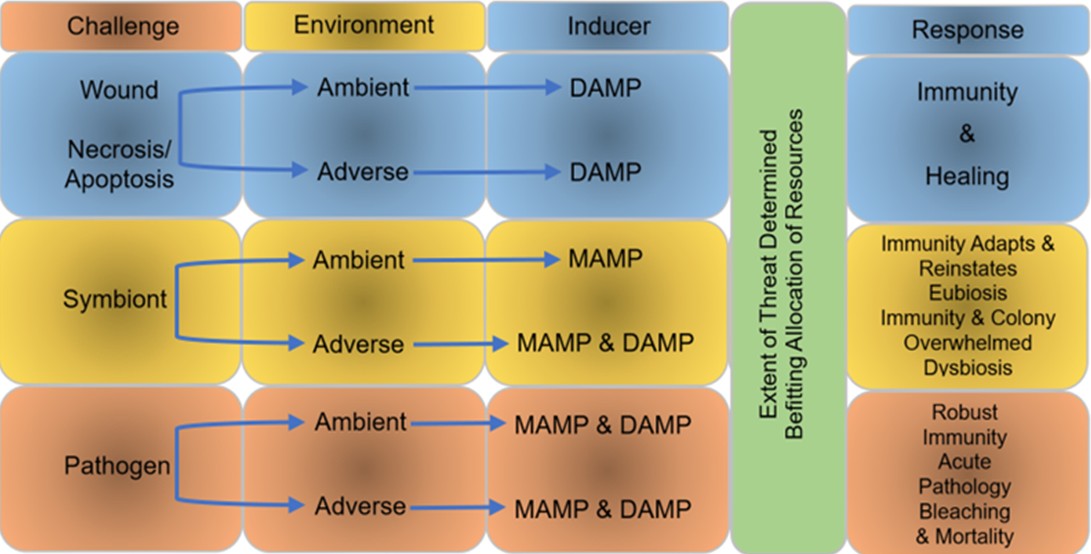
Fig 12. Microbe-associated molecular pattern (MAMP) detection, and/or danger-associated molecular pattern (DAMP) signalling, establish resource allocation and the magnitude of response. Adapted from Palmer 2018.
How can corals maintain remarkably diverse and functionally redundant communities of microbial commensals and symbionts in the face of noxious antiparasitic competencies. Combining the paradigms of self/nonself and the danger model with reference to immune activation may convey an authentic understanding of immune dynamics.
The scale and kind of immune response is likely influenced by holobiont stability and the profile and intensity of signals from microbe- and danger-associated molecular pattern (MAMP; DAMP) receptors. When all is well, MAMP receptors likely detect commensals whilst DAMP receptors remain inactive. When injurious abiotic conditions or pathogens liberate danger-associated molecules from microbes and host cells, the intensity of signals from the holobiont permits the coral to coordinate proportional remediation whilst prudently allocating available resources and reimbursing microbes for consequential harm (Fig 12.). Enlarging this theory provides a dynamic framework whereby corals raise the function of commensals and symbionts whilst kerbing their diversity, moderate pathogens, and navigate acute environmental disruptions (Palmer 2018).
The repertoire of coral defense includes antimicrobial peptides (AMPs) that create holes in the membranes of pathogens and enter to initiate apoptosis (Liu et al. 1995; Vidal-Dupiol et al. 2011; Hlongwane et al. 2018).
The expression of genes associated with immunity, cell endurance, and proliferation are upregulated via the JNK, MAPK p38, and NF-jB pathways after host pattern recognition receptors (PRRs) such as toll-like (TLRs) or nucleotide-binding oligomerisation domain (NOD)-like receptors (NLRs) detect DAMPs. Coral homologues encoding haemolectin and transglutaminase are redolent of sophisticated coagulation mechanisms (Palmer et al. 2012; van de Water et al. 2015). Transglutaminase-mediated plasma protein cross-linking forms an insoluble coagulative gel that hardens into a clot (Palmer et al. 2012). The signalling domains of toll/interleukin receptors (TIRs) are found in multiple proteins associated with innate immunity such as TLR and interleukin-1R receptors (IL-1R), and myeloid differentiation factor 88 (myD88) that may be central to the nurture and shaping of prokaryotic communities (Brown et al. 2023a).
Cellular migration, wound repair, and immunity are amongst numerous downstream mechanisms associated with the activation of receptors known as integrins (Brower et al. 1997; Takada et al. 2007; Kvennefors et al. 2008; Toledo-Hernandez & Ruiz-Diaz 2014, cited in Papke et al. 2024) whose b subunits from the phyla Cnidaria and Porifera are likely to have not changed much over the last five hundred million years (Brower et al. 1997). Expression of integrin-associated genes are upregulated in the cells of Orbicella faveolata in response to SCTLD which includes regenerative integrin alpha-7-like (Lemons & Condic 2008). The cell behavioural and signalling roles of integrins implicates them in the detection of pathogens including those of SCTLD (Juliano et al. 2004; Traylor-Knowles et al. 2021; Pake et al. 2024).

Fig 13. Rescued corals cultured at Apollo Beach’s Florida Aquarium Centre for Conservation (FLAQ). Image courtesy of The Florida Fish and Wildlife Commission ©.
Leucine rich repeats (LRR) are present in certain TLR and toll/interleukin receptors (TIRs; Brown et al. 2023a) like protein 1 and F-box protein 2 of Montastraea cavernosa, protein 15- and 42-like of Orbicella faveolata, and TLR 6 from other Caribbean Scleractinia which are upregulated in those suffering from SCTLD (Traylor-Knowles et al. 2021; Beavers et al. 2023). These kinds of receptor belong to a large family of proteins implicated in pathogen recognition whose downstream cascades help mitigate disease, whereas the b-strand-stabilised folds of LRR, form arcs or horseshoes (Ng & Xavier 2011). LRR contribute to extracellular architectural-complexity which appears to increase sensitivity and “filter” microbial communities (Brown et al. 2023).
DAMP detection induces the melanisation cascade (proPO system) whereby inactive prophenoloxidase (proPO) is cleaved into active phenoloxidase (PO) which expedites the deposition of wound-sealing and pathogen-sequestering melanin (Palmer et al. 2008; Cerenius et al. 2010, cited in Pollock et al. 2019), while highly unstable and toxic quinone intermediates flood and sanitise the lesion (Cerenius et al. 2010, cited in van de Water et al. 2015). Moreover, melanin has antimicrobial properties which may explain the immediate loss of zooxanthellae observed in Porites cylindrica postinjury (Palmer et al. 2011). Unsurprisingly, melanin is abundant in O. faveolata and M. cavernosa suffering from SCTLD (Traylor-Knowles et al. 2021) and may be why lesion-peripheries are strangely darkened in Siderastrea siderea (Fig 7. A).
Fish macrophages internalise cellular debris or dying cells and manufacture the signalling cytokine, tumour necrosis factor-alpha (TNF-α). Transmembrane expressed TNF-α becomes extracellular when cleaved by a metalloprotease, which induces necrosis of atypical cells with pronounced genetic aberrations (Chau et al. 2004; Hong et al. 2013). Rainbow trout’s (Oncorhynchus mykiss’) TNF-α is liberated by several cells including macrophages and T-lymphocytes, whereas the TNF‐β of higher vertebrates appears absent in fish (Laing et al. 2001).

Fig 14. The Marquesas Keys coral rescue cruise where tiny tissue samples from each colony were assayed for genotypic biomarkers. Image courtesy of The Florida Fish and Wildlife Commission ©.
Homologues of TNF and NOD-like receptors together with an arsenal of antimicrobial peptides (AMPs) are synthesised in species of Hydra which are related to Anthozoa that lack scleractinian complement C3 (Miller et al. 2007). Apoptotic genes and those associated with TNF are upregulated in SCTLD-infected corals (Traylor-Knowles et al. 2021; Beavers et al. 2023) which is a typical response to lost tissue (Traylor-Knowles et al. 2021).
“White” cells are common to all animals including corals which phagocytose debris, viruses, and pathogens in phagocytic vesicles which are later infused with digestive enzymes and reactive oxygen species (ROS; Barker et al. 1997; Palmer et al. 2008; Palmer et al. 2011). Phagocytes also kill and sequester pathogens and infected cells with oxidative bursts and form aggregates around foreign invaders, whilst some induce phenoloxidase cascades and melanin deposition.
Fish lectins are essential neutrophil, T-cell, monocyte, and macrophage cell-surface receptors, where lectin dectin-1 is the primary receptor for β-glucans (Brown et al. 2003; Dennehy & Brown 2007; Meena et al. 2012). They are therefore, carbohydrate-binding proteins implicated in cell adhesion which is central to contagion (Endo et al. 2009). Upregulation of the C-type lectin homologue, CTL-2 occurs shortly after abiotic injury in Acropora millepora which soon returns to nominal expression, which suggests its possible participation in the complement system, cellular trauma-specific immunity, and/or the re-establishment of the endosymbiome (van de Water et al. 2015).
MBL-Millectins are receptors that bind zooxanthellae and microbial pathogens that exhibit sequence variation corresponding to their sites of recognition. They caused microbial agglutination in vitro and were upregulated in response to immunostimulation (Kvennefors et al. 2008, cited in van de Water et al. 2015). A Millectin homologue was downregulated postinjury until day seven and increased thereafter to day 10. The gene product Millectin may thus establish and lend stability to the host-zooxanthellae alliance (van de Water et al. 2015).

Fig 15. Florida’s ex situ rescued corals at the Nova Southeastern University in Fort Lauderdale with a comparable oxidation-reduction potential (ORP) to that in the wild. Notwithstanding, the lack of sediment suggests the facility was recently cleansed. Image courtesy of The Florida Fish and Wildlife Commission ©.
After the appearance of tissue loss caused by SCTLD, the expression of genes associated with cell-surface proteins with integral C-type lectin domains are downregulated in Montastraea cavernosa, such as those that bind galactose (Traylor-Knowles et al. 2021). Galactose-binding lectins are key to the adherence of pathogens to the surface of host cells (Elola et al. 2007, cited in Papke et al. 2024; Traylor-Knowles et al. 2021, cited in Papke et al. 2024). Hence the downregulation of cell-surface expressed lectins diminishes infectivity; nevertheless, upregulation of mannose-binding lectins have been associated with the recognition of host-seeking motile mastigote zooxanthellae and pathogens in Orbicella faveolata (Zhou et al. 2017; Traylor-Knowles et al. 2021, cited in Papke et al. 2024).
Intriguingly, necrosis brought on by SCTLD initiates at the base of the endodermal gastrodermis (polyp gut; Landsberg et al. 2020) whereafter zooxanthellae are disrupted (Landsberg et al. 2020; Croquer et al. 2021) before cellular degradation progresses to the ectodermal epidermis (Landsberg et al. 2020).
Dysbiosis and Breakdown of the Host-Zooxanthellae Alliance
Several studies have attempted to elucidate the mechanisms that underpin the bleaching of hermatypic corals that harbour photosynthetic dinoflagellate “algal” symbionts (@zooxanthellae), whose combined efforts perform the swiftest form of biocalcification (Cohen & McConnaughey 2003; Munn 2019). Large polyp stony (LPS) corals appear to regulate and winnow their “algal” partners, the competency of which, may be diminished in small polyp stony (SPS) colonies which turn brown when their zooxanthellae over proliferate (Balling et al. 2008). Moderate and acute bleaching is feasibly restricted to exocytosis in these corals (Fig 16.; Starcevic et al. 2010) while symbionts leave LPS corals via the mesogleal canals and the mesenteries and exudation through the mouth (Rodriguez-Lanetty et al. 2005; Fujise et al. 2014). Alternatively, they may escape from free-floating lysed cells that have lost their contact adhesion (Gates et al. 1992; Boilard et al. 2020; Xu et al. 2023) or become devoid of photoreactive pigments when ROS bleach the thylakoid membranes of their chloroplasts (Boilard et al. 2020; McCauley et al. 2022). Bleaching may be urgent and indiscriminate expulsion of cellular contents (cytosolic “bale out”) in response to a rise in ROS instigated by photosystem overload (Foyer et al. 1994; Fitt & Warner 1995; Müller-Moulé 2002; Starcevic et al. 2010; Fujise et al. 2014). Imported corals appear to dump zooxanthellae, cells, and their contents after days of light deprivation, whereas a recent study demonstrated that the ROS of heat-stressed Scleractinia are unlikely to be generated at a rate that would prove harmful and leak from zooxanthellae (Nielsen et al. 2018). All the same, a “quenching” mechanism in photosystem II of Cladocopium-like Symbiodiniaceae transfers surplus excitation energy to photosystem I, which ceases photosynthetic electron transport and uses the energy to generate heat. The “trigger” may therefore be a consequential loss of photosynthesis and nil exchange of oxygen and carbohydrates (Slavov et al. 2016).
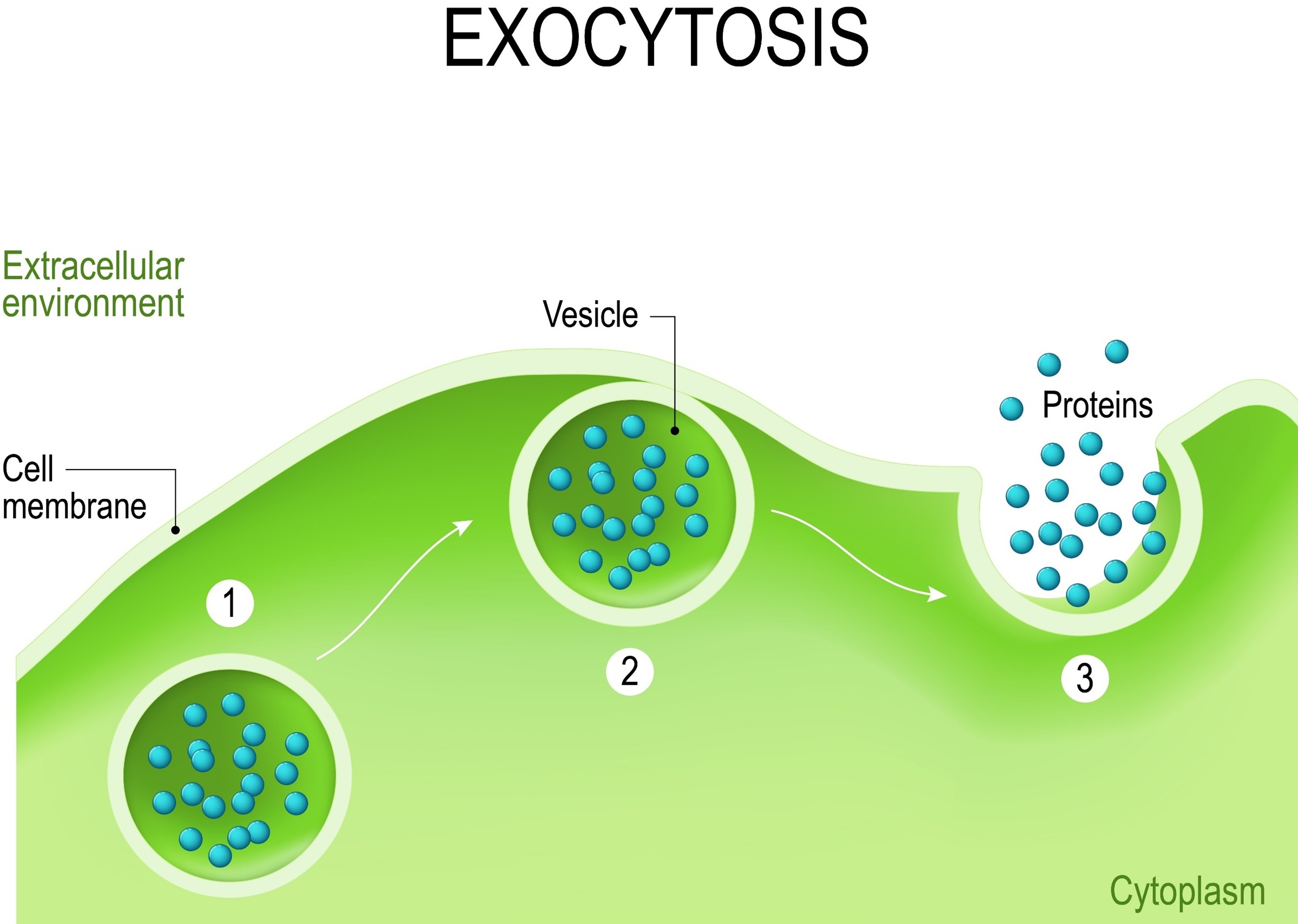
Fig 16. Cells expel waste, and corals may expel symbionts during bleaching using exocytosis (Starcevic et al. 2010) which is distinct from travelling to the mesenteries where they are partially digested before being ejected through the mouth (Rodriguez-Lanetty et al. 2005).
Coccoid “algal” symbionts inhabit membrane-bound organelles called symbiosomes within specialised symbiocytes, which are mesoglea-associated cells of the endodermis (Appendix I; Putnam et al. 2017). Zooxanthellae belong to the family Symbiodiniaceae which transfer photosynthetic oxygen and fixed organic carbon in the form of glucose to their heterotrophic host. Light processing, heat resistance, and photosynthate transference differ amongst genera and strains. Populations are exceedingly diverse where at least clades A to I have been identified (Tonk et al. 2014; Munn 2019). Symbiodinium-, Breviolum-, Cladocopium-, Durusdinium–, Effrenium-, Fugacium-, and Gerakladium-like species were respective clade A, B, C, D, E, Fr5, and G, affiliates. Durusdinium-analogous species confer a profound resistance to bleaching (LaJeunesse et al. 2018).

Fig 17. Recently rescued specimens at the Florida Aquarium Centre for Conservation (FLAQ) at Apollo Beach. Image courtesy of The Florida Fish and Wildlife Commission ©.
Caribbean corals routinely form stable relationships with Symbiodiniaceae of the genera Symbiodinium, Breviolum, Cladocopium, and Durusdinium (Baker 2003; van Oppen & Burghardt 2009, cited in Papke et al. 2024). The findings of numerous field observations, histological studies, and molecular analyses suggest that SCTLD initiates dissolution of symbiosis between the host and its zooxanthellae (Landsberg et al. 2020; Deutsch et al. 2021; Meiling et al. 2021; Work et al. 2021; Studivan et al. 2022b; Williamson et al. 2022; Beavers et al. 2023; Studivan et al. 2023). Despite Beavers and collaborators’ study that demonstrated a statistical correlation between the genus of the predominant in hospite zooxanthellae and the coral’s liability to SCTLD (Beavers et al. 2023), members of the most vulnerable families: Meandrinidae and Mussidae (Baker 2003; Lewis et al. 2018) and other clearly predisposed outgroups (Papke et al. 2024), are home to Breviolum, where some nurture B. psygmophilum (Thornhill et al. 2008, cited in Papke et al. 2024; Leydet & Hellberg 2016, cited in Papke et al. 2024; Lewis et al. 2018). However, this unusual species of former clade B has been discovered in virtually unsusceptible, Cladocora arbuscula and species of Oculina (Papke et al. 2024). Hence robust independent substantiation of the hypothesis of Beavers and collaborators appears lacking.

Fig 18. An ex situ facility conserving specimens from the Florida coral rescue initiative at The National Mississippi River Museum and Aquarium. Image courtesy of The Florida Fish and Wildlife Commission ©.
Other uncertain and somewhat contradictory evidence was acquired in Florida and the U.S. Virgin Islands, where dysbiotic corals that lost pigment appeared to be less vulnerable to SCTLD (Meiling et al. 2020; Sharp et al. 2020; Williams et al. 2021a). Nevertheless, dysbiotic purging of all commensals and symbionts appears to be initiated by the coral, with a view to recruiting populations of pro- and microeu-karyotes better suited to the prevailing conditions (Palmer 2018; Boilard et al. 2020). Forceful clearance of the colony’s mucosal microbiome would also flush out pathogens. Although zooxanthellae are packed full of photoreactive pigments which absorb and fluoresce a myriad of colours, bleaching appears indiscriminate insofar as it may also rid the coral of other potentially useful molecules such as those responsible for pigmentation, because unless associated with wound healing (Palmer et al. 2011) all “algal” symbionts are essentially brown (Appendix I; Yu 2000). Hence mitigation of SCTLD in bleached corals may not necessarily be directly related to a loss of zooxanthellae. We will have to wait until this kind of bioinformatic accrues datasets to perform statistical analyses which may reveal significant trends.
Bacteria: Evidence for Disease Causation
The administration of a combination of antibiotics halts or slows SCTLD (Appendix I; Aeby et al. 2019; Neely et al. 2020; Shilling et al. 2021; Walker et al. 2021; Forrester et al. 2022, cited in Papke et al. 2024), whereas 16 svedberg (S) ribosomal ribonucleic acid (rRNA) next generation sequencing, amplifies and yields the profiles of prokaryotic communities (Woese & Fox 1977; Woese 1987; Weisburg et al. 1991). Numerous studies have used shotgun metagenomics to trace dynamic changes in the microbiomes of corals suffering from SCTLD (Meyer et al. 2019; Rosales et al. 2020; Becker et al. 2021; Evans et al. 2022b; Huntley et al. 2022; Rosales et al. 2022; Studivan et al. 2022b). The findings of this kind of sophisticated analyses have also intimated putative vectors of contagion and spread, such as a sedimental microbial consortium (Studivan et al. 2022b) and biofilms that develop on wet surfaces including the hulls and ballast enclosures of seagoing vessels (Evans et al. 2022a). Members of the bacterial orders Alteromonadales, Campylobacterales, Clostridiales, Flavobacteriales, Peptostreptococcales-Tissierellales, Rhizobiales, Rhodobacterales, and Vibrionales are all comparatively enriched in the lesions of SCTLD compared to the prokaryotic communities of ostensibly healthy corals (Meyer et al. 2019; Rosales et al. 2020; Becker et al. 2021; Clark et al. 2021; Rosales et al. 2022; Rosales et al. 2023). Peptostreptococcales-Tissierellales and Rhodobacterales predominate throughout SCTLD (Rosales et al. 2023) while several transcriptomic and metabolic signatures peculiar to certain microbes, may one day prove useful diagnostic biomarkers (Becker et al. 2021). However, many of these crop up in corals from numerous worldwide regions and thus despite the presence of certain phylotypes and their putative pathogenesis in the lesions of SCTLD, they are unlikely to be the taxa responsible for disease (Papke et al. 2024).

Fig 19. Porcelain disc-mounted propagules. Image courtesy of The Florida Fish and Wildlife Commission ©.
Identification of a microbial agent(s) responsible for SCTLD is exceedingly challenging. Furthermore, advancements in analytical and diagnostic techniques combined with bioinformatics and dataset-appraisals appear to have escalated the enormity of the task. Confoundingly, the microbial communities associated with the lesions of SCTLD vary regionally and in accordance with the age and phase of localised aetiology (Clark et al. 2021; Laas et al. 2021; Rosales et al. 2022; Rosales et al. 2023) as well as the sample site on each specimen (Meyer et al. 2019; Rosales et al. 2020; Clark et al. 2021; Rosales et al. 2022; Rosales et al. 2023).
We do not know if the microorganisms that initiate the disease are the same in every instance, and whether certain lineages that predominate in different kinds of lesion contribute to severity (Aeby et al. 2019). Strains of the notorious coral bleaching bacterium, Vibrio coralliilyticus were present in 20 percent of the acute lesions of M. cavernosa and O. faveolata, which remained essentially absent in those of moderate and chronic pathologies (Ushijima et al. 2020). Strains of this species of bacterium are not the primary pathogens and do not cause the disease but expedite aetiology when present (Papke et al. 2024).
Numerous bacteria undertake the pro-environmental task of digesting and biomineralizing dying or degrading cells and their debris (Egan & Gardiner 2016; Gignoux-Wolfsohn et al. 2017, cited in Papke et al. 2024; Meyer et al. 2019). For instance, saprophytic members of the genus Rhodobacterales appear to opportunistically inhabit the infective boundaries of SCTLD (Rosales et al. 2020).
Diagnostics
SCTLD presents as a lytic necrosis (Papke et al. 2024). Histological mounts facilitate the microscopic examination of ultra-thin tissue cross-sections in a lifelike form. For clarification see: Aslett 2024a. The cellular pathology instigated by SCTLD commences at the skeleton-adjacent calicodermis of the polyp base from where it gradually spreads upwards toward the mouth. Gastrodermal necrosis and mesogleal breakdown is accompanied in some instances by crystalline inclusion bodies (Fig 20. B & D; Landsberg et al. 2020).
Compared to that of basal bodies, necrosis becomes more advanced in upper body walls. The ecto- and endo-dermis, mesoglea, and cell types in the healthy tissues of corals are defined, tightly packed, and precisely arranged, whereas the mesoglea and gastrodermis are disconnected and nebulous, and microscopic anatomies are architecturally necrotised in those suffering from SCTLD (Fig 20.). Rupture of the body wall is preceded by vacuolisation that surrounds symbiocytes and the exocytosis of zooxanthellae (Landsberg et al. 2020; Meiling et al. 2021; Studivan et al. 2022b; Papke et al. 2024).
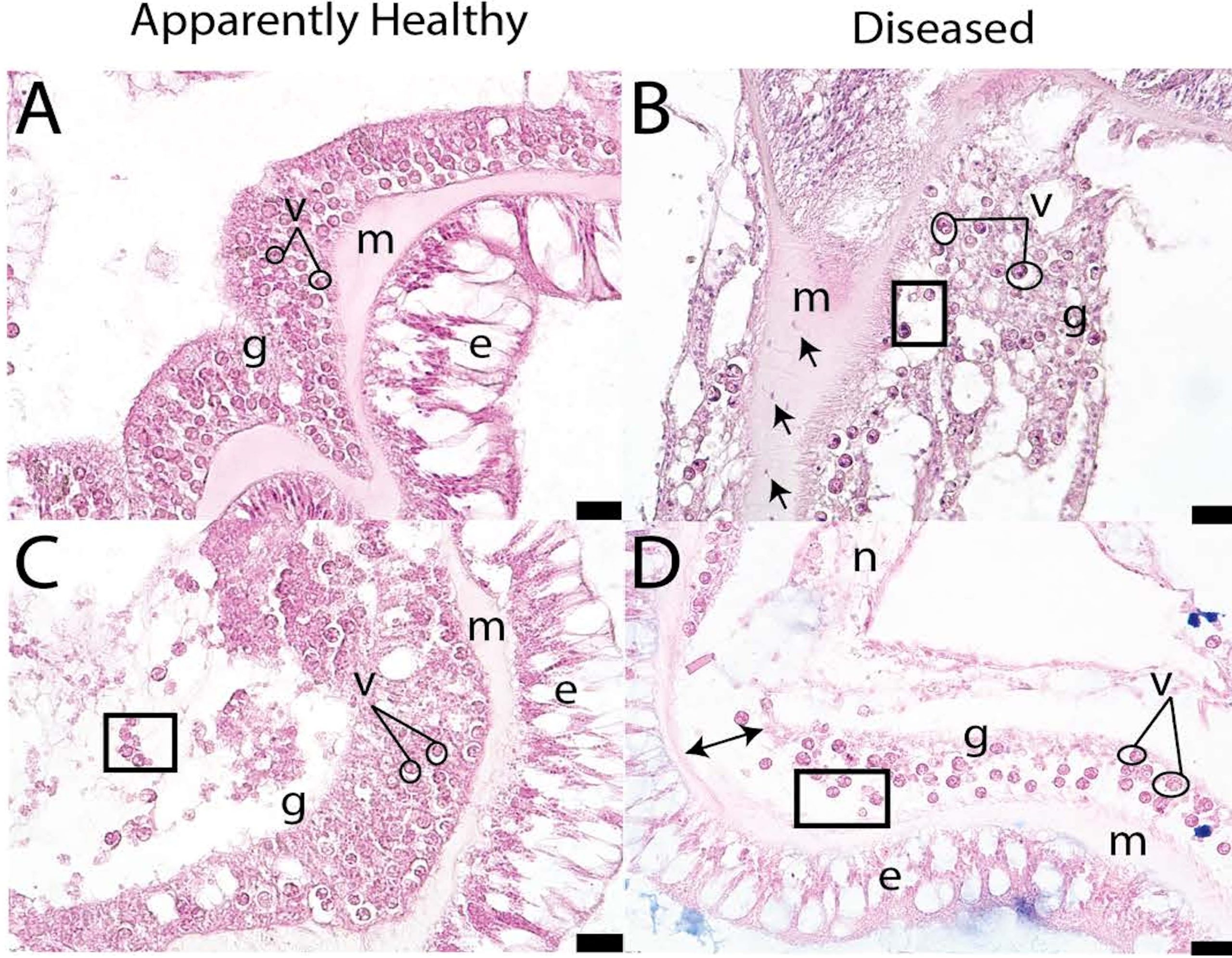
Fig 20. Magnified histologically mounted, stained, and labelled ultrathin cross-sections of the healthy [A & C] and SCTLD-infected [B & D] tissues of Pseudodiploria strigosa. [ם] exocytosis; [e] epidermis; [g] gastrodermis; [↔] separation of the gastrodermis; [→] amoebocytes; [m] mesoglea; [n] necrosis, and [o] vacuolisation. Scale bars 20 µm and adapted from Meiling et al. 2021 and courtesy of Papke et al. 2024 and The Creative Commons Attribution License.
Metabolomic analyses ascertain the abundance of typically small molecules called metabolites and their associated pathways to identify the intracellular mechanisms that underpin the functionality of holobionts and their component organisms. Once a “normal” baseline” is established, comparative studies are performed on diseased or stressed populations to appraise their metabolic responses. Hence these kinds of investigation can estimate wellbeing and identify potential diagnostic biomarkers in asymptomatic carriers or stressed colonies (Lohr et al. 2019; Roach et al. 2020; Little et al. 2021; Roach et al. 2021).
A comparative study of the metabolomic profiles of Floridian M. cavernosa discovered that the metabolites of seemingly healthy colonies are more consistent and stable than those of SCTLD-stricken specimens (Deutsch et al. 2021).
Around one third of all metabolites from healthy and diseased corals were associated with the “algal” endosymbiome, such as diacylglyceryl-carboxyhydroxymethylcholine betaine lipids (Roach et al. 2020; Deutsch et al. 2021; Roach et al. 2021) together with pathways associated with the biosynthesis of vitamin E, glycolipids, and a key component of the thylakoid membranes of zooxanthellae’s chloroplasts (Tchernov et al. 2004; Leblond et al. 2015; Haydon et al. 2023). All things considered, the findings affirmed the “algal” partners of hermatypic corals are affected by the onset of SCTLD (Deutsch et al. 2021).
Since probiotics and beneficial microbes for corals (BMCs; Santoro et al. 2021) appear to redress holobiont dysfunction, metabolomics will likely assist the identification of prokaryotes that produce rare bacteriocins or advantageous metabolites that may be used as part of an inoculum (Papke et al 2024). Metabolomics expedites unearthing the virulence factors of disease-causing agents and possible targets for novel therapeutics.

Fig 21. The in situ culture of a ramet of great star coral (Montastraea cavernosa). Image courtesy of The Florida Fish and Wildlife Commission ©.

Fig 22. Ramet-pruning of relatively scarce staghorn (Acropora cervicornis). Image courtesy of The Florida Fish and Wildlife Commission ©.
Intervention
Preservation of Genetic Homogeneity
The threat of SCTLD motivated the federal agencies of multiple states to bank a vast cross-section of Florida’s corals in 2018 (Schopmeyer, personal communication). The Florida Fish and Wildlife Commission implemented their coral rescue initiative that removed 200 specimens from each of the 20 threatened species from the wild, in order to cultivate them sexually and asexually in public aquaria with their offspring destined to repopulate wild territories. Tiny samples of tissue were taken from each coral which were screened for genetic biomarkers that conferred greater resilience to exogenous a- and -biotic disruptions (Fig 14.). Their genotypes were catalogued and added to a global database, because collecting multiple conspecifics preserves diversity, whilst the population of bioinformatics expedites the identification of resilient strains (Zoccola et al. 2020). It was hoped this strategy would optimise and preserve Florida’s stony coral diversity (Schopmeyer, personal communication).
Similarly, 574 fragments with 128 distinct genotypes of Dendrogyra cylindrus were relocated to in situ nurseries or held in ex situ facilities to prevent the imminent extinction of Floridian examples (Neely et al. 2021b). This dedicated team have rescued 2283 healthy colonies ahead of SCTLD’s advancing front since 2016 whose progress is reported at: https://myfwc.maps.arcgis.com/apps/dashboards/eba7dc2cabc64f60819e6d4b084d94cd.
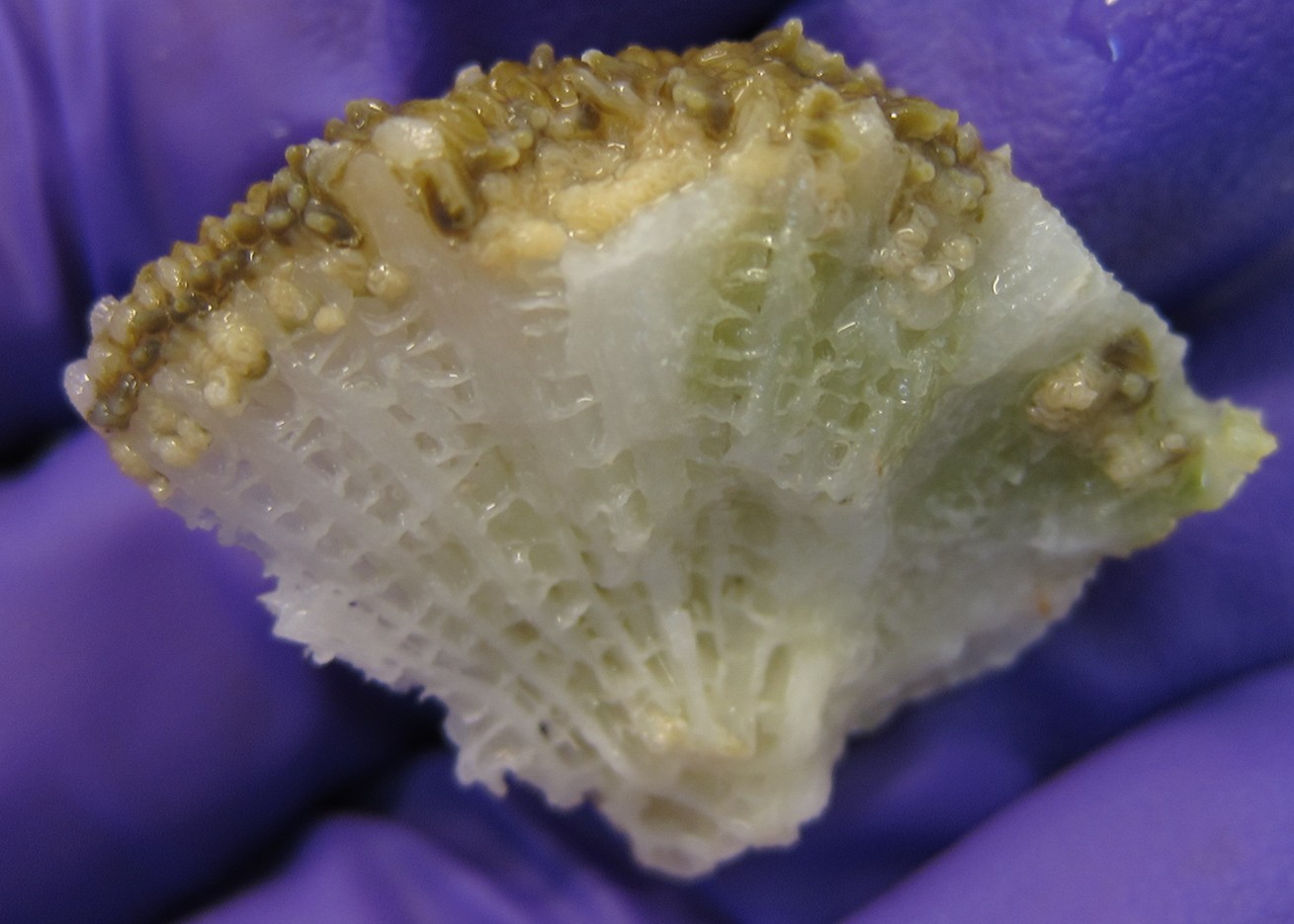
Fig 23. A small fragment of exceedingly susceptible pillar coral, Dendrogyra cylindrus, featuring immature oocytes (Appendix I). Image courtesy of The Florida Fish and Wildlife Commission ©.
Alternatively, and less invasively, cryogenics can freeze coral eggs and spermatozoa (Handwerk 2018). The cryopreserved and thawed sperm from Diploria labyrinthiformis which suffer acute SCTLD, successfully fertilised recently harvested eggs (Grosso-Becerra et al. 2021).
Colonies could be rehomed in remote oligotrophic oceanic locations, which is ordinarily unworkable because suitably qualified people require shelter, sustenance, healthcare, and technology. Notwithstanding, evidence suggests that corals relocated vast distances from their geographic origins may not adapt to their new environment (Berger, personal communication). Why do therefore, most imports survive in our artificial reefs. Science raises questions and tends to guard against obvious contradictions, but we are not infallible. Consequently, the success of aquarists would infer that accurate appraisals and protracted monitoring of the local ecology at the candidate location would improve survivorship. Consult: The Complete Reef Aquarist – A Conservation Manual for further guidance (https://www.ebay.co.uk/itm/116390701502).
Amputation
The diseased parts of a coral can be physically separated from the healthy residuum. Corals respond well to swift decisive forceful excisions with sharp chisels and other brutal instruments. Like in captivity, amputation is invariably unsuccessful possibly due to invasive stress, and besides, whether in the wild or a recirculating system, the pathogen remains present and thus reinfects and annihilates the colony. It simply delays the inevitable, whilst ex situ eutrophy proves significantly injurious. Thus were the outcomes of detaching part colonies or relocating pillars of in situ Dendrogyra cylindrus (Neely & Hower 2019, cited in Papke et al. 2024).

Fig 24. In situ photographs of SCTLD-infected corals off leeward Bonaire: [A] Pseudodiploria strigosa; [B] Diploria labyrinthiformis; [C] Colpophyllia natans; [D] Eusmilia fastigiata; [E] Meandrina meandrites, and [F] Montastraea cavernosa. Courtesy of Pepe et al. 2025 and The Creative Commons Attribution License.
Akin to the mandatory ruthless instant elimination of degrading or dying specimens in recirculating systems, removing infected colonies in the wild to prevent the spread of a disease would conceivably prove efficacious (Johnston 2021). Culling would have to be implemented shortly after a disease outbreak with the aim of forestalling an epizootic (Bolzoni et al. 2014). By virtue of SCTLD’s foothold in the Caribbean, it is too late to use this tactic there (Precht 2021). This strategy might prove useful during, heaven forbid, the emergence of SCTLD in hitherto uninfected worldwide regions, whilst retrieving boulders is not for the fainthearted (Papke et al. 2024).
Trenching and Antibiotics
Creating a 1 cm deep firebreak 5 cm from the advancing lesion in combination with amoxicillin, halted 91 percent of lesions and mitigated the destruction of 86 percent of colonies (Appendix I; Shilling et al. 2021; Walker et al. 2021), where just firebreaks were less effective at 79 and 38 percent (Shilling et al. 2021). However, this research corroborates we are dealing with a bacterium or a consortium thereof; and attests the efficacy of amoxicillin whose resistance is likely encoded on conjugable plasmids that are ubiquitously disseminated amongst bacteria (Sodhi et al. 2021). Perhaps the prokaryotes from the lesions of SCTLD could be screened for genes that confer amoxicillin resistance to reveal a subpopulation of candidate causative agents. Trenching was considered unworkable thanks to its laboriousness and somewhat ineffectiveness (Shilling et al. 2021).
Chlorinated Epoxy
A blend of two-part marine epoxy and calcium hypochlorite otherwise known as chlorine powder, is applied to the lesion periphery (Aeby et al. 2015; Neely et al. 2021c; Shilling et al. 2021; Walker et al. 2021). Personal protective equipment (PPE) such as gloves, eye protection, and a face visor and mask are mandatory, as epoxy is toxic and chlorine powder is exceedingly corrosive. Amalgamate in a well-ventilated area. Despite some success mitigating black band disease (Aeby et al. 2015), this strategy proves merely marginally beneficial (Lee Hing et al. 2022), insofar as the rates of arresting disease were akin to those where the illness spontaneously abates (Neely et al. 2021c; Shilling et al. 2021; Walker et al. 2021).

Fig 25. Detaching a small colony from a reef using a stone mallet and cold chisel. Image courtesy of The Florida Fish and Wildlife Commission ©.
Antibiotics
Amoxicillin, ampicillin, chloramphenicol, kanamycin, and sulphathiazole retain varying degrees of efficacy (Aeby et al. 2019; Miller et al. 2020), whilst ampicillin and kanamycin appear to act synergistically, yet they exert variable outcomes when used on their own. Amoxicillin appears to be the b-lactam antibiotic of choice, that has expedited 97 percent survival of ex situ fragments (Miller et al. 2020).
Ocean Alchemist supply the wave action resistant coral mucus-simulating ointment, CoralCureTM base2B. When combined with antibiotics, it slowly releases them over three days. It is administered directly to the boundary of infection with 60-ml syringes as it does not adhere to living tissue (Neely et al. 2020; Neely et al. 2021c), or it may be impregnated into rope (Gay 2021; Eaton et al. 2022). It was not devised for aquaria because antibiotics negatively impact beneficial microbes including the biofilter consortium.
When CoralCureTM was mixed with amoxicillin trihydrate and applied to infected Floridian C. natans, D. labyrinthiformis, D. stokesii, M. cavernosa, O. annularis, O. faveolata, P. clivosa, P. strigosa, and S. siderea, remission was observed in 88 to 95 percent of colonies (Neely et al. 2021c; Shilling et al. 2021; Walker et al. 2021; Lee Hing et al. 2022), whereas trials in the British Virgin Islands, were redolent of curtailing spread (Neely et al. 2021c, cited in Papke et al. 2024; Forrester et al. 2022, cited in Papke et al. 2024). Remarkably, like the effects of beneficial microbes for corals (BMCs; Santoro et al. 2021), amoxicillin restores cross-anatomy gene expression to that of undiseased colonies (Studivan et al. 2023).
Although this strategy was the most successful and preferred in 2024, it is laborious, expensive, and does not prevent the development of new lesions, and thus relies upon frequent reapplications (Neely et al. 2021c; Shilling et al. 2021; Walker et al. 2021; Lee Hing et al. 2022). Furthermore, the extensive use of antibiotics not only predisposes microbes to resistance but has a widescale impact on reef ecology, because conventional water-purifying chemolithoautotrophs may be killed along with vital mucosal populations.

Fig 26. An in situ ramet of rare Acropora cervicornis where surprisingly, over 90 percent were thriving after three months. Image courtesy of The Florida Fish and Wildlife Commission ©.
Chemotherapy
Iron is essential for all life where higher eukaryotes typically obtain it from their diet. However, prokaryotes require inorganic dissolved iron (DIFe2+/DIFe3+) and thus many liberate iron-scavenging siderophores which are later internalised to release their payload. Lesion-boundary-proximal iron sequestration has been proposed as a potential therapy. Combining the iron chelator, Chelex 100 and 2-bipyridyl (BIP) with coral dental paste was ineffective ex situ. However, the researchers remarked that the synthetic salt supplier had begun to add an iron-containing anticaking agent which may have confounded analyses (Miller et al. 2020). The immune systems of humans sequester iron as an antibacterial maneuver; however, no matter how challenging its efficacy is to estimate, people still get virulent and fatal bacterial infections.
Beneficial Microbes for the Mucosal Microbiome
Varied mucosal communities are vital for colony defense where community shifts or a change in diversity, may represent host-mediated dysbiotic purging in response to pathogenesis or dysfunction (Rosenberg et al. 2007; Munn 2019). Probiotics can therefore raise performance and restore health (Rosado et al. 2018; Munn 2019; Santoro et al. 2021).

Fig 27. A sizeable colony of Eusmilia fastigiata integral to the Florida coral rescue initiative mounted with reef putty on a terracotta tile. Image courtesy of The Florida Fish and Wildlife Commission ©.

Fig 28. A Floridian coral rescue initiative brain coral (Colpophyllia natans). Image courtesy of The Florida Fish and Wildlife Commission ©.
The microbiome flexibility, probiotic, and beneficial microorganisms for corals (BMCs) hypotheses propose that the plasticity of a microbiome differs amongst species; the composition of a coral holobiont is optimised to suit the prevailing conditions which remains susceptible to probiotic manipulation, and resilience to stress is linked to symbiotic populations such as the consortium within mucus. By these tokens, microbial inoculums can mitigate bleaching and disease (Reshef et al. 2006; Peixoto et al. 2017; Rosado et al. 2018; Munn 2019; Voolstra & Ziegler 2020; Santoro et al. 2021).
Probiotics applied to the mucosal microbiomes of heat stressed corals restructured their microbial communities and instigated genetic and metabolic alterations that ameliorated bleaching and mortality. Beneficial microbes for corals (BMCs) applied to thermally stressed hermatypes elicited holobiont reorganisation and modification of cross-anatomy gene expression to favour recuperative cellular repair, immunity, and thermal protection. Furthermore, the revitalising bacterial consortium proved a biocide for Vibrio coralliilyticus strain V1 and Vibrio alginolyticus V2 (Santoro et al. 2021).
The BMC isolates were sourced from healthy corals, albeit inexplicably, the mucosal consortia of the probiotic-treated colonies did not reflect the profile of the inoculum, while the communities of those that were not compromised did not alter when exposed. However, the transcriptome of each coral coordinated with the profile of its mucosal microbiome, which implies coral immunity and the holobiont communicate via receptors and molecular exchange (Santoro et al. 2021). Such findings corroborate crosstalk and hologenomics (Rosenberg & Zilber-Rosenberg 2018), whereas recuperation coincided with the reversion of each mucosal microbiome to its pre-stressed profile (Santoro et al. 2021).
Probiotics
Most people are familiar with human dietary probiotics where live cultures of “good” bacteria are purchased and ingested. “Good” bacteria enhance the performance of mucosal microbiomes insofar as they promote the health of the underlying cells like enterocytes or gill and skin epithelia, which assists the upkeep of an effective barrier. Diverse populations of beneficial prokaryotes outcompete their harmful counterparts and impede their colonisation. They also quorum quench and manufacture rare and useful metabolites that promote the wellbeing of the holobiont, or they may be exceedingly efficient sequestrators of inorganic iron.
Amongst the conventional repertoire of antibacterial enzymes, prokaryotes of the strain McH1-7 of the genus Pseudoalteromonas produce the biocides: koromicin and tetrabromopyrrole which arrested the progression of SCTLD in 59 percent of cases. Most significantly, it forestalled transmission ex situ when it was exploited as a prophylactic inoculum (Ushijima et al. 2023). This probiotic may thus be a viable contender for shielding in situ colonies (Papke et al. 2024).
Mucosal microbiota are the first line of colony defence, whose destabilisation might lead to extensive localised decimation. The mass culture and inoculation of the wild may therefore exert unpredictable effects. Even so, in situ bacteriophage therapy of coral disease has proven to slow progression and lessen contagion of coral plague on Red Sea Dipsastraea favus (Atad et al. 2012, cited in Papke et al. 2024; Garber 2012), which was not a new idea (Aslett 2003; Efrony et al. 2007; Efrony et al. 2009; Teplitski & Ritchie 2009). Reassuringly, the functional mucosal communities of ostensibly healthy colonies appear to retain an inertia, insofar as they remained unchanged when inoculated with BMCs (Santoro et al. 2021).
SCTLD appears to raise more questions than it answers. Its persistence and competency for rapidly extending its range into adjacent geographic territories feasibly abetted by shipping, poses the most profound threat to Caribbean and conceivably worldwide reef ecology thus far.
Appendix I
Photographs I & II: exceedingly rare glimpses of bundle broadcasting in healthy elkhorn (Acropora palmata) and oocyte broadcasting from a maternal colony of pillar coral (Dendrogyra cylindrus; Fig 23.) courtesy of The Florida Fish and Wildlife Commission ©. Photograph III: the coccoid in hospite form of Symbiodinium “fitti” found in Caribbean acroporids and giant clams. Acropora cervicornis, A. palmata, and their hybrid, A. prolifera enjoy an enduring association with S. “fitti” type A3 (Pinzón et al. 2010; Reich et al. 2021). Image, Todd C. LaJeunesse ©. Photography IV: [column I] the SCTLD advancing front specified by white arrows; [column II] firebreak indicated by the black arrow with the lesion boundary treated with antibiotic paste, and [column III] the cured and healed coral a month later. Courtesy of Walker et al. 2021 and The Creative Commons Attribution License. To demonstrate how challenging it is to correctly identify an illness from nothing but observation, could photograph V be an image of Acropora cervicornis with SCTLD or A. muricata suffering from something else in another part of the world. Photograph VI: elkhorn.





References
Abbas, M., N., Kausar, S. & Cui, H. (2019) The biological role of peroxiredoxins in innate immune responses of aquatic invertebrates. Fish Shellfish Immunol. 89, 91-97. doi: 10.1016/j.fsi.2019.03.062
Aebu, G., S. & Santavy, D., L. (2006) Factors affecting susceptibility of the coral Montastraea faveolata to black-band disease. Marine Ecology Progress Series. Inter-Research, Luhe, Germany. 318,103-110.
Aeby, G., S. et al. (2019) Pathogenesis of a tissue loss disease affecting multiple species of corals along the Florida Reef Tract. Front. Mar. Sci. 6, 1-18.
Aeby, G., Ushijima, B., Bartels, E., Walter, C., Kuehl, J., Jones, S. et al. (2021) Changing stony coral tissue loss disease dynamics through time in montastraea cavernosa. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.699075
Ainsworth, T., Kvennefors, E., Blackall, L., Fine, M. & Hoegh-Guldberg, O. (2007) Disease and cell death in White Syndrome of Acroporid corals on the Great Barrier Reef. Marine Biology. 151(1),.
Alvarez-Filip, L., Estrada-Saldívar, N., Pérez-Cervantes, E., Molina-Hernández, A. & González-Barrios, F., J., A (2019) Rapid spread of the Stony Coral Tissue Loss Disease outbreak in the Mexican Caribbean. PeerJ. https://doi.org/10.7717/peerj.8069
Alvarez-Filip, L., González-Barrios, F., J., Pérez-Cervantes, E., Molina-Hernández, A. & Estrada-Saldívar, N. (2022) Stony coral tissue loss disease decimated Caribbean coral populations and reshaped reef functionality. Communications Biology. 5, 440. 440 https://doi.org/10.1038/s42003-022-03398-6; www.nature.com/commsbio
Ambalavanan, L., Iehata, S., Fletcher, R., Stevens, E., H. & Zainathan, S., C. (2021) A review of marine viruses in coral ecosystem. J. Mar. Sci. Eng. 9, 711. doi: 10.3390/ jmse9070711
Aronson, R., B. & Precht, W., F. (2001) White-band disease and the changing face of Caribbean coral reefs. Hydrobiologia. 460, 25-38.
Arrigoni, R., Benzoni, F., Huang, D., Fukami, H., Chen, C., A., Berumen, M., L., Hoogenboom, M., Thomson, D., P., Hoeksema, B., W., Budd, A., F., Zayasu, Y., Terraneo, T., I., Kitano, Y., F. & Baird, A., H. (2016) When forms meet genes: revision of the scleractinian genera Micromussa and Homophyllia (Lobophylliidae) with a description of two new species and one new genus. Contributions to Zoology. 85, 387-422.
Aslett, C., G. (2003) Bacteriophage Therapy of Coral Disease. Master of Research thesis, The University of Plymouth, UK.
Aslett, C., G. (2024) Holosystemics Parts I to VIII. https://www.reefranch.co.uk/
Aslett, C., G. (2024a) Reef Focus: Microscopes and Histology: An Introduction. https://www.reefranch.co.uk/
Baker, A., C. (2003) Flexibility and specificity in coral-algal symbiosis: diversity, ecology, and biogeography of symbiodinium. Annu. Rev. Ecology Evolution Systematics. 34, 661-689. doi: 10.1146/annurev.ecolsys.34.011802.132417
Barker, L., P., George, K., M., Falkow, S. & Small, P., L. (1997) Differential trafficking of live and dead Mycobacterium marinum organisms in macrophages. Infection and immunity. 65(4), 1497-1504.
Bass, D., Stentiford. G., D., Wang, H-C., Koskella, B. & Tyler, C., R. (2019) The pathobiome in animal and plant diseases. Trends Ecol Evol. 34(11), 996-1008.
Beavers, K., M., Van Buren, E., W., Rossin, A., M., Emery, M., A., Veglia, A., J., Karrick, C., E. et al. (2023) Stony coral tissue loss disease induces transcriptional signatures of in situ degradation of dysfunctional Symbiodiniaceae. Nat. Commun. 14, 2915. doi: 10.1038/s41467-023-38612-4
Becker, C., C., Brandt, M., Miller, C., A. & Apprill, A. (2021) Microbial bioindicators of Stony Coral Tissue Loss Disease identified in corals and overlying waters using a rapid field-based sequencing approach. Environ. Microbiol. 24 (3), 1166-1182. doi: 10.1111/1462-2920.15718
Becker, C., C., Weber, L., Zgliczynski, B., Sullivan, C., Sandin, S., Muller, E. et al. (2023) Microorganisms and dissolved metabolites distinguish Florida’s Coral Reef habitats. PNAS Nexus. 2, pgad287. doi: 10.1093/pnasnexus/pgad287
Berger, M. The University of Leeds, UK.
Berthelier, J., Schnitzler, C., Wood-Charlson, E., Poole, A., Weis, V. & Detournay, O. (2017) Implication of the host TGFβ pathway in the onset of symbiosis between larvae of the coral Fungia scutaria and the dinoflagellate Symbiodinium sp. (clade C1f). Coral Reefs. 36, 1-6.
Birrell, C., Mccook, L., Willis, B. & Diaz-Pulido, G. (2008) Effects of benthic algae on the replenishment of corals and the implications for the resilience of coral reefs. Oceanography and marine biology. 46,.
Boilard, A., Dubé, C., E., Gruet, C., Mercière, A., Hernandez-Agreda, A. & Derome, N. (2020) Defining Coral Bleaching as a Microbial Dysbiosis within the Coral Holobiont. Microorganisms. 8, 1682. https://doi.org.10.3390/microorganisms8111682
Bolzoni, L., Tessoni, V., Groppi, M. & De Leo, G. (2014) React or wait: which optimal culling strategy to control infectious diseases in wildlife. J. Math Biol. 69, 1001-1025. doi: 10.1007/s00285-013-0726-y
Borneman, E. (2002) Coralmania: The Food of Reefs, Part 3: Phytoplankton. ReefKeeping.com. http://www.reefkeeping.com/issues/2002-10/eb/index.php
Brandt, M., E., Ennis, R., S., Meiling, S., S., Townsend, J., Cobleigh, K., Glahn, A. et al. (2021) The emergence and initial impact of stony coral tissue loss disease (SCTLD) in the United States Virgin islands. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.7153
Brower, D., L., Brower, S., M., Hayward, D., C. & Ball, E., E. (1997). Molecular evolution of integrins: Genes encoding integrin b subunits from a coral and a sponge. Proc. Natl. Acad. Sci. U.S.A. 94, 9182-9187. doi: 10.1073/pnas.94.17.9182
Brown, G., D., Herre, J., Williams, D., L., Willment, J., A., Marshall, A., S., J. & Gordon, S. (2003) Dectin-1 Mediates the Biological Effects of β-Glucans. The Journal of Experimental Medicine. 197(9), 1119-1124.
Brown, T., Sonett, D., McMinds, R., Pollock, F., J., Medina, M. & Zaneveld, J., R. (2023) Anatomically-specific coupling between innate immune gene repertoire and microbiome structure during coral evolution. https://doi.org/10.1101/2023.04.26.538298
Brown, T., Sonett, D., McMinds, R., Pollock, F., J., Medina, M. & Zaneveld, J., R. (2023a) Anatomically-specific coupling between innate immune gene repertoire and microbiome structure during coral evolution. https://doi.org/10.1101/2023.04.26.538298
Bulleri, F., Thiault, L., Mills, S., C., Nugues, M., Eckert, E., Corno, G. & Joachim, C. (2018) Erect macroalgae influence epilithic bacterial assemblages and reduce coral recruitment. Marine Ecology Progress Series. 597, 65-77.
Bythell, J., Pantos, O. & Richardson, L. (2004) White Plague, White Band, and Other “White” Diseases. Coral Health and Disease. Rosenberg, E. & Loya, Y. (eds.). Springer, Berlin, Heidelberg. pp 351-365.
Camacho-Vite, C., Estrada-Saldıvar, N., Perez-Cervantes, E. & Alvarez-Filip, L. (2022) Differences in the progression rate of SCTLD in Pseudodiploria strigosa are related to colony size and morphology. Front. Mar. Sci. 9, 790818. doi: 10.3389/ fmars.2022.790818
Cardenas, A., Rodriguez-R, L., M., Pizarro, V., Cadavid, L., F. & Arevalo-Ferro, C. (2011) Shifts in bacterial communities of two caribbean reef-building coral species affected by white plague disease. The Isme Journal. 6, 502.
Casamatta, D., Stanic, D., Gantar, M. & Richardson, L. (2012) Characterization of Roseofilum reptotaenium (Oscillatoriales, Cyanobacteria) gen. et sp. nov. isolated from Caribbean Black band disease. Phycologia. 51(5), 489.
Cerenius, L., Kawabata, S., Lee, B., L., Nonaka, M. & Soderhall, K. (2010) Proteolytic cascades and their involvement in invertebrate immunity. Trends Biochem. Sci. 35, 575-583. https://doi:10.1016/j.tibs. 2010.04.006
Chau, B., N., Chen, T., T., Wan, Y., Y., DeGregori, J. & Wang, J., Y. (2004) Tumor necrosis factor alpha-induced apoptosis requires p73 and c-ABL activation downstream of RB degradation. Molecular and cellular biology. 24(10), 4438-4447.
Chaudhury, A. & Howe, P., H. (2009) The tale of transforming growth factor-beta (TGFbeta) signaling: a soigné enigma. IUBMB life. 61(10), 929-939.
Clark, A., S., Williams, S., D., Maxwell, K., Rosales, S., M., Huebner, L., K., Landsberg, J., H. et al. (2021) Characterization of the microbiome of corals with stony coral tissue loss disease along Florida’s coral reef. Microorganisms. 9, 2181. doi: 10.3390/ microorganisms9112181
Clemens, E. & Brandt, M., E. (2015) Multiple mechanisms of transmission of the Caribbean coral disease white plague. Coral Reefs. 34, 1179-1188. doi: 10.1007/s00338 015-1327-6
Combs, I., R., Studivan, M., S., Eckert, R., J. & Voss, J., D. (2021) Quantifying impacts of stony coral tissue loss disease on corals in Southeast Florida through surveys and 3D photogrammetry. PloS One. 16, e0252593. doi: 10.1371/journal.pone.0252593
Cooney, R., P., Pantos, O., Tissier, M., D., Barer, M., R., O´Donnell, A., G. & Bythell, J., C. (2002) Characterization of the bacterial consortium associated with black band disease in coral using molecular microbiological techniques. Environmental Microbiology. 4, 401-413.
Costa, S., V., Hibberts, S., J., Olive, D., A., Budd, K., A., Long, A., E., Meiling, S., S. et al. (2021) Diversity and disease: the effects of coral diversity on prevalence and impacts of stony coral tissue loss disease in Saint Thomas, U.S. Virgin Islands. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.682688
Croquer, A., Weil, E. & Rogers, C., S. (2021) Similarities and differences between two deadly Caribbean coral diseases: white plague and stony coral tissue loss disease. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.709544
De Bakker, D., M., Meesters, E., H., Bak, R., P., M., Nieuwland, G. & Van Duyl, F., C. (2019) Long-term shifts in coral communities on shallow to deep reef slopes of curacao and bonaire: The rise of octocorals as major reef builders. Front. Mar. Sci. 6, 66. doi: 10.3389/fmars.2019.00066
De Bakker, D., M., Meesters, H., Bak, R., P., M., Nieuwland, G. & Van Duyl, F., C. (2016) Long-term shifts in coral communities on shallow to deep reef slopes of Curacao and Bonaire: Are there any winners? Front. Mar. Sci. 3. doi: 10.3389/ fmars.2016.00247
De Bakker, D., M., Van Duyl, F., C., Bak, R., P., M., Nugues, M., M., Nieuwland, G. & Meesters, H. (2017) 40 Years of benthic community change on the Caribbean reefs of Curacao and Bonaire: the rise of slimy cyanobacterial mats. Coral Reefs 36, 355-367. doi: 10.1007/s00338-016-1534-9
Dennehy, K., M. & Brown, G., D. (2007) The role of the β‐glucan receptor Dectin‐1 in control of fungal infection. Journal of Leukocyte Biology. 82, 253-258.
Deutsch, J., M., Jaiyesimi, O., A., Pitts, K., A., Houk, J., Ushijima, B., Walker, B., K. et al. (2021) Metabolomics of healthy and stony coral tissue loss disease affected montastraea cavernosa corals. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.714778
Deutsch, J., M., Mandelare-Ruiz, P., Yang, Y., Foster, G., Routhu, A., Houk, J. et al. (2022) Metabolomics approaches to dereplicate natural products from coral-derived bioactive bacteria. J. Nat. Prod. 85, 462-478. doi: 10.1021/acs.jnatprod.1c01110
Dobbelaere, T., Holstein, D., M., Muller, E., M., Gramer, L., J., McEachron, L., Williams, S., D. et al. (2022) Connecting the dots: transmission of stony coral tissue loss disease from the marquesas to the Dry Tortugas. Front. Mar. Sci. 9. doi: 10.3389/ fmars.2022.778938
Dobbelaere, T., Muller, E., M., Gramer, L., J., Holstein, D., M. & Hanert, E. (2020) Coupled epidemio-hydrodynamic modeling to understand the spread of a deadly coral disease in Florida. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.591881
Eaton, K., R., Clark, A., S., Curtis, K., Favero, M., Hanna Holloway, N., Ewen, K. & Muller, E., M. (2022) A highly effective therapeutic ointment for treating corals with black band disease. PLoS ONE. 17(10), e0276902.
Eaton, K., R., Landsberg, J., H., Kiryu, Y., Peters, E., C. & Muller, E., M. (2021) Measuring Stony Coral Tissue Loss Disease Induction and Lesion Progression Within Two Intermediately Susceptible Species, Montastraea cavernosa and Orbicella faveolata. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.717265
Efrony, R., Atad, I. & Rosenberg, E. (2009) Phage Therapy of Coral White Plague Disease: Properties of Phage BA3. Curr Microbiol. 58, 139.
Efrony, R., Loya, Y., Bacharach, E. & Rosenberg, E. (2007) Phage therapy of coral disease. Coral Reefs. 26, 7.
Egan, S. & Gardiner, M. (2016) Microbial dysbiosis: rethinking disease in marine ecosystems. Front. Microbiol. 7. doi: 10.3389/fmicb.2016.00991
Elmer, F., Cruz, Y., Dock, C., Ortega, A., Wedel, S. & Hertler, H. (2021) Determining lesion progression rate of stony coral tissue loss disease using structure from-motion photogrammetry. GCR. 32, SC12-SC17. doi: 10.18785/gcr.3201.15
Endo, Y., Nakazawa, N., Iwaki, D., Takahashi, M., Matsushita, M. & Fujita, T. (2009) Interactions of ficolin and mannose-binding lectin with fibrinogen/fibrin augment the lectin complement pathway. J. Innate Immun. 2, 33-42. doi: 10.1159/ 000227805
Estrada-Saldıvar, N., Molina-Hernandez, A., Perez-Cervantes, E., Medellın Maldonado, F., Gonzalez-Barrios, F., J. & Alvarez-Filip, L. (2020) Reef-scale impacts of the stony coral tissue loss disease outbreak. Coral Reefs. 39, 861–866. doi: 10.1007/s00338-020-01949-z
Estrada-Saldívar, N., Quiroga-García, B., A., Pérez-Cervantes, E., Rivera-Garibay, O., O. & Alvarez-Filip, L. (2021) Effects of the stony coral tissue loss disease outbreak on coral communities and the benthic composition of cozumel reefs. Front. Mar. Sci. 8, 1-13.
Evans, J., S., Paul, V., J. & Kellogg, C. A. (2022a) Biofilms as potential reservoirs of stony coral tissue loss disease. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.1009407
Evans, J., S., Paul, V., J., Ushijima, B. & Kellogg, C., A. (2022b) Combining tangential flow filtration and size fractionation of mesocosm water as a method for the investigation of waterborne coral diseases. Biol. Methods Protoc. 7, bpac007. doi: 10.1093/biomethods/bpac007
Evans, J., S., Paul, V., J., Ushijima, B., Pitts, K., A. & Kellogg, C., A. (2023) Investigating microbial size classes associated with the transmission of stony coral tissue loss disease (SCTLD). PeerJ. 11, e15836. doi: 10.7717/peerj.15836
Fitt, W., K. & Warner, M., E. (1995) Bleaching Patterns of Four Species of Caribbean Reef Corals. The Biological Bulletin. 189(3), 298-307.
Forrester, G., E., Arton, L., Horton, A., Nickles, K. & Forrester, L., M. (2022) Antibiotic treatment ameliorates the impact of stony coral tissue loss disease (SCTLD) on coral communities. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.859740
Foyer, C., H., Lelandais, M. & Kunert, K., J. (1994) Photooxidative stress in plants. Physiologia Plantarum. 92, 696-717.
Fuess, L., E., Butler, C., C., Brandt, M., E. & Mydlarz, L., D. (2020) Investigating the roles of transforming growth factor-beta in immune response of Orbicella faveolata, a scleractinian coral. Developmental and comparative immunology. 107, 103639.
Fuess, L., E., Pinz, J., H., Weil, E. & Mydlarz, L., D. (2016) Associations between transcriptional changes and protein phenotypes provide insights into immune regulation in corals. Dev. Comp. Immunol. 62, 17-28. https://doi.org/10.1016/j.dci.2016.04.017
Fujise, L., Yamashita, HY., Suzuki, G., Sasaki, K., Liao, L., M. & Koike, K. (2014) Moderate Thermal Stress Causes Active and Immediate Expulsion of Photosynthetically Damage Zooxanthellae (Symbiodinium) from Corals. PLOS ONE. DOI:10.1371/journal.pone.0114321
Garber, S. (2012) Favia coral receives virus therapy for white plague disease. AdvancedAquarist.com. https://www.advancedaquarist.com/blog/favia-coral-receives-phage-therapy-for-white-plague-like-disease
Gates, R., D., Baghdasarian, G. & Muscatine, L. (1992) Temperature stress causes host cell detachment in symbiotic cnidarians: implications for coral bleach-ing. Biol Bull. 182, 324–332. https://doi.org/10.2307/1542252
Gay, J. (2021) CoralCure Ointment Base2B is a topical treatment for Stony Coral Tissue Loss Disease. ReefBuilders.com. https://reefbuilders.com/2021/11/10/coralcure-ointment-base2b-is-a-topical-treatment-for-stony-coral-tissue-loss-disease/
Giambiagi-Marval, M., Mafra, M., A., Penido, E., G. & Bastos, M., C. (1990) Distinct groups of plasmids correlated with bacteriocin production in Staphylococcus aureus. Journal of general microbiology. 136(8), 1591-1599.
Gignoux-Wolfsohn, S., A., Aronson, F., M. & Vollmer, S., V. (2017) Complex interactions between potentially pathogenic, opportunistic, and resident bacteria emerge during infection on a reef-building coral. FEMS Microbiol. Ecol. 93. doi: 10.1093/femsec/fix080
Godar, D. (1996) Preprogrammed and Programmed Cell Death Mechanisms of Apoptosis: UV‐Induced Immediate and Delayed Apoptosis. Photochemistry & Photobiology. 63(6),.
Goreau, T., Cervino, J., Goreau, M., Hayes, R., Hayes, M., Richardson, L., Smith, G., DeMeyer, K., Nagelkerken, I., Garzon-Ferreira, J., Gil-Agudelo, D., Peters, E., Garrison, G., Williams, E., Jr., Williams, L., Quirolo, C., Patterson, K., Porter, J., W. & Porter, K. (1998) Rapid spread of diseases in Caribbean coral reefs. Revista de Biologia Tropica. 46, 157-172.
Green, E., P. & Bruckner, A., W. (2000) The significance of coral disease epizootiology for coral reef conservation. Biol. Conserv. 96, 347-361. doi: 10.1016/S0006-3207(00)00073-2
Grosso-Becerra, M., V., Mendoza-Quiroz, S., Maldonado, E. & Banaszak, A., T. (2021) Cryopreservation of sperm from the brain coral Diploria labyrinthiformis as a strategy to face the loss of corals in the Caribbean. Coral Reefs 40, 937-950. doi: 10.1007/s00338-021-02098-7
Guzman-Urieta, E., O. & Jordan-Dahlgren, E. (2021) Spatial patterns of a lethal white syndrome outbreak in pseudodiploria strigosa. Front. Mar. Sci. 8. doi: 10.3389/ fmars.2021.669171
Hall, E., R., Muller, E., M., Goulet, T., Bellworthy, J., Ritchie, K., B. & Fine, M. (2018) Eutrophication may compromise the resilience of the Red Sea coral Stylophora pistillata to global change. Marine Pollution Bulletin. 131, 701-711.
Halliwell, B. & Gutteridge, J., M., C. (2015) Free Radicals in Biology and Medicine. Oxford University Press, Oxford, United Kingdom. doi: 10.1093/acprof:oso/ 9780198717478.001.0001
Handwerk, B. (2018) Coral Larvae Cryogenically Frozen and Thawed for the First Time. SmithsonianMag.com. https://www.smithsonianmag.com/science-nature/coral-larvae-cryogenically-frozen-and-thawed-first-time-180970905/
Harvell, C., D. et al. (2007) Coral diseases, Environmental drivers and the balance between corals and microbial associates. Oceanography. 20, 172-195.
Haydon, T., D., Matthews, J., L., Seymour, J., R., Raina, J-B., Seymour, J., E., Chartrand, K. et al. (2023) Metabolomic signatures of corals thriving across extreme reef habitats reveal strategies of heat stress tolerance. Proc. R. Soc. B: Biol. Sci. 290, 20221877. doi: 10.1098/rspb.2022.1877
Hayes, N., K., Walton, C., J. & Gilliam, D., S. (2022) Tissue loss disease outbreak significantly alters the Southeast Florida stony coral assemblage. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.975894
Heres, M., M., Farmer, B., H., Elmer, F. & Hertler, H. (2021) Ecological consequences of stony coral tissue loss disease in the Turks and Caicos Islands. Coral Reefs. 40, 609-624. doi: 10.1007/s00338-021-02071-4
Hlongwane, P., Mungra, N., Madheswaran, S., Akinrinmade, O., A., Chetty, S. & Barth, S. (2018) Human Granzyme B Based Targeted Cytolytic Fusion Proteins. Biomedicines. 6(2), 72.
Ho, L. (2013) A newly discovered bacterial denitrification symbiosis. AdvancedAquarist.com. https://www.advancedaquarist.com/blog/a-newly-discovered-bacterial-denitrification-symbiosis
Hogarth, W., T. (2006) Endangered and threatened species: final listing determinations for elkhorn and staghorn coral. Fed. Regist. 71, 26852-26861.
Hong, S., Li, R., Xu, Q., Secombes, C., J. & Wang, T. (2013) Two Types of TNF-α Exist in Teleost Fish: Phylogeny, Expression, and Bioactivity Analysis of Type-II TNF-α 3 in Rainbow Trout Oncorhynchus mykiss. The Journal of Immunology. 191(12), 5959-5972.
Howe-Kerr, L., I., Knochel, A., M., Meyer, M., D., Sims, J., A., Karrick, C., E., Grupstra, C., G., B. et al. (2023) Filamentous virus-like particles are present in coral dinoflagellates across genera and ocean basins. ISME J. 17, 1-14. doi: 10.1038/ s41396-023-01526-6
Huang D (2012) Threatened Reef Corals of the World. PLoS ONE 7(3): e34459. doi:10.1371/journal.pone.0034459
Huang, Z., Langevelde, F., Estrada-Peña, A., Suzan, G. & de Boer, W., F. (2016) The diversity-disease relationship: Evidence for and criticisms of the dilution effect. Parasitology 1, 1-12. doi: 10.1017/S0031182016000536
Hubbard, S., R. & Till, J., H. (2000) Protein tyrosine kinase structure and function. Annual review of biochemistry. 69, 373-398.
Huntley, N., Brandt, M., E., Becker, C., C., Miller, C., A., Meiling, S., S., Correa, A., M., S. et al. (2022) Experimental transmission of Stony Coral Tissue Loss Disease results in differential microbial responses within coral mucus and tissue. ISME Commun. 2, 46. doi: 10.1038/s43705-022-00126-3
International Coral Reef Initiative (2019) Stony coral tissue loss disease. https://www.icriforum.org/news/2019/04/stony-coral-tissue-loss-disease
Jeon, H-S., Dracheva, T., Yang, S-H., Meerzaman, D., Fukuoka, J., Shakoori, A. et al. (2008) SMAD6 contributes to patient survival in non–small cell lung cancer and its knockdown reestablishes TGF-b Homeostasis in lung cancer cells. Cancer Res. 68, 9686-9692. doi: 10.1158/0008-5472.CAN-08-1083
Jin, Y., K., Lundgren, P., Lutz, A., Raina, J., B., Howells, E., J., Paley, A., S., Willis, B., L. & van Oppen, M., J. (2016) Genetic markers for antioxidant capacity in a reef-building coral. Science advances. 2(5), e1500842.
Johnston, M., A. (2021) Strategy for Stony Coral Tissue Loss Disease Prevention and Response at Flower Garden Banks National Marine Sanctuary. Department of Commerce, National Oceanic and Atmospheric Administration, Galveston, TX.
Joyner, J., L., Sutherland, K., P., Kemp, D., W., Berry, B., Griffin, A., Porte, J., W., Amador, M., H., Noren, H., K. & Lipp, E., K. (2015) Systematic Analysis of White Pox Disease in Acropora palmata of the Florida Keys and Role of Serratia marcescens. Appl Environ Microbiol. 81(13), 4451-4457.
Juliano, R., L., Reddig, P., Alahari, S., Edin, M., Howe, A. & Aplin, A. (2004) Integrin regulation of cell signalling and motility. Biochem. Soc. Trans. 32 (3), 443-446. doi: 10.1042/bst0320443
Keesing, F., Holt, R., D. & Ostfeld, R., S. (2006). Effects of species diversity on disease risk. Ecol. Lett. 9, 485-498. doi: 10.1111/j.1461-0248.2006.00885.x
Kolodziej, G., Studivan, M., S., Gleason, A., C., R., Langdon, C., Enochs, I., C. & Manzello, D., P. (2021) Impacts of stony coral tissue loss disease (SCTLD) on coral community structure at an inshore patch reef of the upper Florida keys using photomosaics. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.682163
Kramer, P., R., Roth, L. & Lang, J. (2020) Map of Coral Cover of Susceptible Coral Species to SCTLD. ArcGIS Online. www.agrra.org.
Kvennefors, E., C., E., Leggat, W., Hoegh-Guldberg, O., Degnan, B., M. & Barnes, A., C. (2008) An ancient and variable mannose-binding lectin from the coral Acropora millepora binds both pathogens and symbionts. Dev. Comp. Immunol. 32, 1582-1592. doi: 10.1016/j.dci.2008.05.010
Kvennefors, E., C., E., Leggat, W., Kerr, C., C., Ainsworth., T., D., Hoegh-Guldberg., O. & Barnes, A., C. (2010) Analysis of evolutionarily conserved innate immune components in coral links immu- nity and symbiosis. Developmental and Comparative Immunology. 34, 1219-1229.
Laas, P., Ugarelli, K., Absten, M., Boyer, B., Briceño, H. & Stingl, U. (2021) Composition of prokaryotic and eukaryotic microbial communities in waters around the Florida reef tract. Microorganisms. 9, 1120. doi: 10.3390/microorganisms9061120
Laing, K., J., Wang, T., Zou, J., Holland, J., Hong, S., Bols, N., Hirono, I., Aoki, T. & Secombes, C., J. (2001) Cloning and expression analysis of rainbow trout Oncorhynchus mykiss tumour necrosis factor‐α. European Journal of Biochemistry. 268, 1315-1322.
LaJeunesse, T., C., Parkinson, J., E., Gabrielson, P., W., Jeong, H., J., Reimer, J., D., Voolstra, C., R. & Santos, S., R. (2018) Systematic Revision of Symbiodiniaceae Highlights the Antiquity and Diversity of Coral Endosymbionts. Current biology: CB. 28(16), 2570-2580.
LaJeunesse, T., Parkinson, J., E. & Trench, R., K. (2012) Symbiodinium. http://tolweb.org/Symbiodinium/126705
Landsberg, J., H. et al. (2020) Stony coral tissue loss disease in Florida is associated with disruption of Host–Zooxanthellae Physiology. Front. Mar. Sci. 7, 1-24.
Landsberg, J., H., Kiryu, Y., Peters, E., C., Wilson, P., W., Perry, N., Waters, Y. et al. (2020) Stony coral tissue loss disease in Florida is associated with disruption of host zooxanthellae physiology. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.576013
Lavy, A., Keren, R., Yu, K., Thomas, B., C., Alvarez-Cohen, L., Banfield, J., F. & Ilan, M. (2018) A novel Chromatiales bacterium is a potential sulfide oxidizer in multiple orders of marine sponges. Environmental microbiology. 20(2), 800-814.
Leblond, J., D., Khadka, M., Duong, L. & Dahmen, J., L. (2015) Squishy lipids: Temperature effects on the betaine and galactolipid profiles of a C18/C18 peridinin containing dinoflagellate, Symbiodinium microadriaticum (Dinophyceae), isolated from the mangrove jellyfish, Cassiopea xamachana. Phycological Res. 63, 219-230. doi: 10.1111/pre.12093
Lee Hing, C., Guifarro, Z., Dueñas, D., Ochoa, G., Nunez, A., Forman, K. et al. (2022) Management responses in Belize and Honduras, as stony coral tissue loss disease expands its prevalence in the Mesoamerican reef. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.883062
Lemons, M., L. & Condic, M., L. (2008) Integrin signaling is integral to regeneration. Exp. Neurol. 209, 343-352. doi: 10.1016/j.expneurol.2007.05.027
Lesser, M., P., Falcón, L., I., Rodríguez-Román, A., Enríquez, S., Hoegh-Guldberg, O. & Iglesias-Prieto, R. (2007) Nitrogen fixation by symbiotic cyanobacteria provides a source of nitrogen for the scleractinian coral Montastraea cavernosa. Mar Ecol Prog Ser. 346, 143-152.
Lessios, H., A. (2016) The great diadema antillarum die-off: 30 years later. Annu. Rev. Mar. Sci. 8, 267-283. doi: 10.1146/annurev-marine-122414-033857
Lessios, H., A., Robertson, D., R. & Cubit, J., D. (1984) Spread of Diadema mass mortality through the Caribbean. Science. 226, 335-337.
Lewis, A., M., Chan, A., N. & LaJeunesse, T., C. (2018) New species of closely related endosymbiotic dinoflagellates in the greater Caribbean have niches corresponding to host coral phylogeny. J. Eukaryot. Microbiol. 66, 469-482. doi: 10.1111/jeu.12692
Leydet, K., P. & Hellberg, M., E. (2016) Discordant coral–symbiont structuring: factors shaping geographical variation of Symbiodinium communities in a facultative zooxanthellate coral genus, Oculina. Coral Reefs. 35, 583-595. doi: 10.1007/s00338-016 1409-0
Little, M., George, E., E., Arts, M., G., I., Shivak, J., Benler, S., Huckeba, J. et al. (2021) Three-dimensional molecular cartography of the Caribbean reef-building coral orbicella faveolata. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.627724
Liu, C., C., Walsh, C., M. & Young, J., D., E. (1995) Perforin: structure and function. Immunology Today. 16(4), 194-201.
Lohr, K., E., Khattri, R., B., Guingab-Cagmat, J., Camp, E., F., Merritt, M., E., Garrett, T., J. et al. (2019) Metabolomic profiles differ among unique genotypes of a threatened Caribbean coral. Sci. Rep. 9, 6067. doi: 10.1038/s41598-019-42434-0
Maire, J. & van Oppen, M., J., H. (2022) A role for bacterial experimental evolution in coral bleaching mitigation? Trends in microbiology. 30(3), 217-228.
McCauley, M., Goulet, T., L., Jackson, C., R. & Loesgen, S. (2022) Systematic review of cnidarian microbiomes reveals insights into the structure, specificity, and fidelity of marine associations. Nature Communications. 14, 4899. https://doi.org/10.1038/s41467-023-39876-6
Meena, D., Das, P., Kumar, S., Mandal, S., Prusty, A., Singh, S., Akhtar, M., Behera, B., Kumar, K., Pal, A. & Mukherjee, S. (2012) Beta-glucan: an ideal immunostimulant in aquaculture (a review). Fish Physiology and Biochemistry. 39(3), 431-457.
Meiling, S., Muller, E., M., Smith, T., B., and Brandt, M., E. (2020) 3D photogrammetry reveals dynamics of stony coral tissue loss disease (SCTLD) lesion progression across a thermal stress event. Front. Mar. Sci. 7. doi: 10.3389/ fmars.2020.597643
Meiling, S., S., Muller, E., M., Lasseigne, D., Rossin, A., Veglia, A. J., MacKnight, N., et al. (2021) Variable species responses to experimental stony coral tissue loss disease (SCTLD) exposure. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.670829
Meyer, J., L., Castellanos-Gell, J., Aeby, G., S., Häse, C., C., Ushijima, B. & Paul, V., J. (2019) Microbial community shifts associated with the ongoing stony coral tissue loss disease outbreak on the Florida reef tract. Front. Microbiol. 10. doi: 10.3389/ fmicb.2019.02244
Michael, J., K., Cameron V., J. & Manuel, N. (2011) Attenuation coefficients of ultraviolet and photosynthetically active wavelengths in the waters of Heron Reef, Great Barrier Reef, Australia. Marine & Freshwater Research. 63(2), 142-149.
Miller, A., W. & Richardson, L., L. (2015) Emerging coral diseases: a temperature driven process? Mar. Ecol. 36, 278-291. doi: 10.1111/maec.12142
Miller, C., May, L., Moffitt, Z. & Woodley, C. (2020) “Exploratory treatments for stony coral tissue loss disease : pillar coral (Dendrogyra cylindrus),” in NOAA Technical Memorandum NOS NCCOS 245 NOAA and CRCP. 37. 78-78.
Miller, D., J., Hemmrich, G., Ball, E., E., Hayward, D., C., Khalturin, K., Funayama, N. et al. (2007) The innate immune repertoire in Cnidaria- ancestral complexity and stochastic gene loss. Genome Biol. 8, R59. doi: 10.1186/gb-2007-8-4-r59
Miller, M., W., Lohr, K., E., Cameron, C., M., Williams, D., E. & Peters, E., C. (2014) Disease dynamics and potential mitigation among restored and wild staghorn coral, Acropora cervicornis. PeerJ 2, e541. doi: 10.7717/peerj.541
Molina-Hernández, A., González-Barrios, F., J., Perry, C., T. & Álvarez-Filip, L. (2020) Two decades of carbonate budget change on shifted coral reef assemblages: Are these reefs being locked into low net budget states? Caribbean reefs carbonate budget trends. Proc. Royal Soc. B: Biol. Sci. 287. https://doi.org/10.1098/rspb.2020.2305rspb20202305
Morrow, K., M., Muller E. & Lesser M., P. (2018) How Does the Coral Microbiome Cause, Respond to, or Modulate the Bleaching Process? Coral Bleaching. Ecological Studies (Analysis and Synthesis). 233. van Oppen, M. & Lough, J. (eds.). Springer, Cham. pp 153-188.
Muller, E., M., Sartor, C., Alcaraz, N., I. & van Woesik, R. (2020) Spatial epidemiology of the stony-coral-tissue-loss disease in Florida. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00163
Müller-Moulé, P., A. (2002) Ascorbate Deficiency Can Limit Violaxanthin De-Epoxidase Activity in Vivo. Coral disease. 128(3), 970.
Munn, C., B. (2019) Marine Microbiology: Ecology & Applications, Third Edition. Munn, C., B. (ed.). CRC Press, Taylor & Francis Group, London. pp 273-326.
NCBI (2019) The National Center for Biotechnology Information, Taxonomy Browser. https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi
Neely, K. & Hower, E. (2019) FY 2018 In Situ Disease Intervention. Florida Department of Environmental Protection, Miami, FL.
Neely, K., L., Lewis, C., L., Lunz, K., S. & Kabay, L. (2021a) Rapid population decline of the pillar coral dendrogyra cylindrus along the Florida reef tract. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.656515
Neely, K., L., Lewis, C., L., O’Neil, K., Woodley, C., M., Moore, J., Ransom, Z. et al. (2021b) Saving the last unicorns: the genetic rescue of Florida’s pillar corals. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.657429
Neely, K., L., Macaulay, K., A., Hower, E., K. & Dobler, M., A. (2020) Effectiveness of topical antibiotics in treating corals affected by Stony Coral Tissue Loss Disease. PeerJ. doi: 10.7717/peerj.9289
Neely, K., L., Shea, C., P., Macaulay, K., A., Hower, E., K. & Dobler, M., A. (2021c) Short- and long-term effectiveness of coral disease treatments. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.675349
Ng, A. & Xavier, R., J. (2011) Leucine-rich repeat (LRR) proteins: Integrators of pattern recognition and signaling in immunity. Autophagy 7, 1082-1084. doi: 10.4161/ auto.7.9.16464
Nguyen-Kim, H., Bettarel, Y., Bouvier, T., Bouvier, C., Doan-Nhu, H., Nguyen-Ngoc, L. et al. (2015) Coral mucus is a hot spot for viral infections. Appl. Environ. Microbiol. 81, 5773-5783. doi: 10.1128/AEM.00542-15
Nielsen, D., A., Petrou, K. & Gates, R., D. (2018) Coral bleaching from a single cell perspective The ISME Journal. 12, 1558–1567. https://doi.org/10.1038/s41396-018-0080-6
Noonan, K., R. & Childress, M., J. (2020) Association of butterflyfishes and stony coral tissue loss disease in the Florida Keys. Coral Reefs. 39, 1581-1590. doi: 10.1007/ s00338-020-01986-8
Palmer, C., V. & Traylor-Knowles, N. (2012) Towards an integrated network of coral immune mechanisms. Proc. R. Soc. B: Biol. Sci. 279, 4106-4114. doi: 10.1098/ rspb.2012.1477
Palmer, C., V. (2018) Immunity and the coral crisis. Communications Biology. https://dio.org/10.1038/s42003-018-0097-4
Palmer, C., V., Bythell, J., C. & Willis, B., L. (2010) Levels of immunity parameters underpin bleaching and disease susceptibility of reef corals. The FASEB Journal. 24,.
Palmer, C., V., Bythell, J., C. & Willis, B., L. (2012) Enzyme activity demonstrates multiple pathways of innate immunity in Indo-Pacific anthozoans. Proceedings of the Royal Society B: Biological Sciences. 279, 3879-3887.
Palmer, C., V., McGinty, E., S., Cummings, D., J., Smith, S., M., Bartels, E. & Mydlarz, L., D. (2011a) Patterns of coral ecological immunology: variation in the responses of Caribbean corals to elevated temperature and a pathogen elicitor. J. Exp. Biol. 214, 4240-4249. doi: 10.1242/jeb.061267
Palmer, C., V., Mydlarz, L., D. & Willis, B., L. (2008) Evidence of an inflammatory-like response in non-normally pigmented tis- sues of two scleractinian corals. Proceedings of the Royal Society B: Biological Sciences. 275, 2687-2693.
Palmer, C., V., Traylor-Knowles, N., G., Willis, B., L. & Bythell, J., C. (2011) Corals use similar immune cells and wound-healing processes as those of higher organisms. PLoS ONE. 6, e23992.
Pantos, O., Cooney, R., Le Tissier, M., Barer, M., O’Donnell, A. & Bythell, J. (2003). The bacterial ecology of a plague‐like disease affecting the Caribbean coral Montastrea annularis. Environmental Microbiology. 5(5), 370-382.
Papke, E., Carreiro, A., Dennison, C., Deutsch, J., M., Isma, L., M., Meiling, S., S., Rossin, A., M., Baker, A., C., Brandt, M., E., Garg, N., Holstein, D., M., Traylor-Knowles, N., Voss, J., D. & Ushijima, B. (2024) Stony coral tissue loss disease: a review of emergence, impacts, etiology, diagnostics, and intervention. Front. Mar. Sci. 10:1321271. doi: 10.3389/fmars.2023.1321271
Peixoto, R., S., Rosado, P., M., Leite, D., C., Rosado, A., S. & Bourne, D., G. (2017) Beneficial Microorganisms for Corals (BMC): Proposed Mechanisms for Coral Health and Resilience. Frontiers in microbiology. 8, 341.
Penczykowski, R., M., Laine, A., L. & Koskella, B. (2015) Understanding the ecology and evolution of host-parasite interactions across scales. Evolutionary applications. 9(1), 37-52.
Pepe, B., A., Van der Roest, V., Vlam, O., Nouse, S. & Meesters, E. (2025) Assessing the impact of stony coral tissue loss disease on coral cover on Bonaire’s Leeward side. Front. Mar. Sci. 11, 1512371. doi: 10.3389/fmars.2024.1512371
Perry, C., T. & Alvarez-Filip, L. (2019) Changing geo-ecological functions of coral reefs in the Anthropocene. Funct. Ecol. 33, 976-988.
Perry, C., T. et al. (2014) Changing dynamics of Caribbean reef carbonate budgets: emergence of reef bioeroders as critical controls on present and future reef growth potential. Proc. R. Soc. B: Biol. Sci. 281, 20142018-20142018.
Pinzón, J., Devlin-Durante, M., Weber, M., Baums, I. & LaJeunesse, T. (2010) Microsatellite loci for Symbiodinium A3 (S. fitti) a common algal symbiont among Caribbean Acropora (stony corals) and Indo-Pacific giant clams (Tridacna). Conservation Genetics Resources. 3(1), 45-47.
Pollock, F., J., Lamb, J., B., van de Water, J., Smith, H., A., Schaffelke, B., Willis, B., L. & Bourne, D., G. (2019) Reduced diversity and stability of coral-associated bacterial communities and suppressed immune function precedes disease onset in corals. R Soc Open Sci. 6, 190355. https://doi.org/10.1098/rsos .190355
Pollock, F.,J., McMinds, R., Smith, S., Bourne, D., G., Willis, B., L., Medina, M., Thurber, R., V. & Zaneveld, J., R. (2018) Coral-associated bacteria demonstrate phylosymbiosis and cophylogeny. Nat Commun. 9, 4921.
Porter, J., W., Dustan, P., Japp, W., Patterson, K., L., Kosmynin, V., Meier, O., W., Patterson, M., E. & Parsons, M., Ò. (2001) Patterns of spread of coral disease in the Florida Keys. Hydrobiologia. 460, 1-24.
Precht, W., F., Gintert, B., E., Robbart, M., L., Fura, R. & van Woesik, R. (2016) Unprecedented disease-related coral mortality in southeastern Florida. Sci. Rep. 6, 31374. doi: 10.1038/srep31374
Preston, E. (2015) Deadly “White Syndrome” Kills Coral, and it’s Spreading. Hakai Magazine, Costal Science and Societies. https://www.hakaimagazine.com/news/deadly-white-syndrome-kills-coral-and-its-spreading/
Proceedings of the National Academy of Sciences, 201812412. https://www.pnas.org/content/early/2019/02/05/1812412116.full
Purdy, E. & Winterer, E. (2005) Contradicting Barrier Reef relationships for Darwin’s Evolution of reef types. International Journal of Earth Sciences. 95(1), 143-167.
Putnam, H., Barott, K., Ainsworth, T. & Gates, R. (2017) The Vulnerability and Resilience of Reef-Building Corals. Current Biology. 27(11), 1551-1706.
Quiroz, S., M., Renteria, R., T., Tapia, G., G., R., Miller, M., W., Grosso-Becerra, M., V. & Banaszak, A., T. (2023) Coral affected by stony coral tissue loss disease can produce viable offspring. PeerJ. 11, e15519. doi: 10.7717/peerj.15519
Ramirez-Portilla, C., Baird, A., H., Cowman3, P., F., Quattrini, A., M., Harii, S., Frederic Sinniger, F. & Flot, J-F. (2022) Solving the Coral Species Delimitation Conundrum. Syst. Biol. 71(2), 461-475. http://www.dio.org/10.1093/sysbio/syab077
Reich, H., Kitchen, S., Stankiewicz, K., Devlin‐Durante, M., Fogarty, N. & Baums, I. (2021) Genomic variation of an endosymbiotic dinoflagellate (Symbiodinium ‘fitti’) among closely related coral hosts. Molecular Ecology. 30(14), 3500-3514.
Reshef, L., Koren, O., Loya, Y., Zilber-Rosenberg, I. & Rosenberg, E. (2006) The coral probiotic hypothesis. Environmental microbiology. 8(12), 2068-2073.
Richardson, L., L., Goldberg, W., M., Carlton, R., G. & Halas, J., C. (1998) Coral disease outbreak in the Florida Keys: Plague Type II. Revista de Biologia Tropica. 46, 187-198.
Richardson, L., L., Sekar, R., Myers, J., L., Gantar, M., Voss, J., D., Kaczmarsky, L., Remily, E., R., Boyer, G., L. & Zimba, P., V. (2007) The presence of the cyanobacterial toxin microcystin in black band disease of corals. FEMS Microbiology Letters. 272(2), 182-187.
Ritchie, K., B. & Smith, W., G. (1998) Type II white-band disease. Revista de Biologia Tropica. 46, 199-203.
Roach, T., N., F., Dilworth, J., H., C., M., Jones, A., D., Quinn, R., A. & Drury, C. (2021) Metabolomic signatures of coral bleaching history. Nat. Ecol. Evol. 5, 495–503. doi: 10.1038/s41559-020-01388-7
Roach, T., N., F., Little, M., Arts, M., G., I., Huckeba, J., Haas, A., F., George, E., E. et al. (2020) A multiomic analysis of in situ coral–turf algal interactions. Proc. Natl. Acad. Sci. 117, 13588-13595. doi: 10.1073/pnas.1915455117
Robbins, S., J., Singleton, C., M., Chan, C., X., Messer, L., F., Geers, A., U., Ying, H., Baker, A., Bell, S., C., Morrow, K., M., Ragan, M., A., Miller, D., J., Forêt, S., ReFuGe2020 Consortium, Voolstra, C., R., Tyson, G., W. & Bourne, D., G. (2019) A genomic view of the reef-building coral Porites lutea and its microbial symbionts. Nature microbiology. 4(12), 2090-2100.
Rodriguez-Lanetty, M., Scaramuzzi, C., Quinnell, R. & Larkum, A. (2005) Transport of symbiotic zooxanthellae in mesogleal canals of Zoanthus robustus? Coral Reefs. 24, 195-196.
Rosado, P., Leite, D., Duarte, G., Chaloub, R., Jospin, G., Nunes da Rocha, U., P., Saraiva, J., Dini-Andreote, F., Eisen, J., Bourne, D. & Peixoto, R. (2018) Marine probiotics: increasing coral resistance to bleaching through microbiome manipulation. The ISME Journal. 13(4), 921-936.
Rosales, S., M., Clark, A., S., Huebner, L., K., Ruzicka, R., R., and Muller, E., M. (2020) Rhodobacterales and rhizobiales are associated with stony coral tissue loss disease and its suspected sources of transmission. Front. Microbiol. 11. doi: 10.3389/ fmicb.2020.00681
Rosales, S., M., Huebner, L., K., Clark, A., S., McMinds, R., Ruzicka, R., R. & Muller, E., M. (2022) Bacterial metabolic potential and micro-eukaryotes enriched in stony coral tissue loss disease lesions. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.776859
Rosales, S., M., Huebner, L., K., Evans, J., S., Apprill, A., Baker, A., C., Becker, C., C. et al. (2023) A meta-analysis of the stony coral tissue loss disease microbiome finds key bacteria in unaffected and lesion tissue in diseased colonies. ISME Commun. 3, 1-14. doi: 10.1038/s43705-023-00220-0
Rosenberg, E. & Zilber-Rosenberg, I. (2018) The hologenome concept of evolution after 10 years. Microbiome. 6(1), 78.
Rosset, S., L., Koster, G., Brandsma, J., Hunt, A., N., Postle, A., D. & D’Angelo, C. (2019) Lipidome analysis of Symbiodiniaceae reveals possible mechanisms of heat stress tolerance in reef coral symbionts. Coral Reefs 38, 1241-1253. doi: 10.1007/ s00338-019-01865-x
Santoro, E., P., Borges, R., M., Espinoza, J., L., Freire, M., Messias, C., S., M., A., Villela, H., D., M., Pereira, L., M., Vilela, C., L., S., Rosado, J., G., Cardoso, P., M., Rosado, P., M., Assis, J., M., Duarte, G., A., S., Perna, G., Rosado, A., S., Macrae, A., Dupont, C., L., Nelson, K., E., Sweet, M., J., Voolstra, C., R. & Peixoto, R., S. (2021) Coral microbiome manipulation elicits metabolic and genetic restructuring to mitigate heat stress and evade mortality. Science advances. 7(33), 3088.
Schmitt, R., J., Holbrook, S., J., Davis, S., L., Brooks, A., J. & Adam, T., C. (2019) Experimental support for alternative attractors on coral reefs.
Schopmeyer, S., A. (2021) Associate Research Scientist. Fish and Wildlife Research Institute. St. Petersburg, FL 33701.
Sharp, W., C. & Maxwell, K., E. (2018) Investigating the ongoing coral disease outbreak in the Florida Keys: Collecting corals to diagnose the etiological agent(s) and establishing sentinel sites to monitor transmission rates and the spatial progression of the disease. Final Report to the Florida Department of Environmental Protection.
Sharp, W., C., Shea, C., P., Maxwell, K., E., Muller, E., M. Hunt, J., H. (2020) Evaluating the small-scale epidemiology of the stony-coral-tissue-loss-disease in the middle Florida keys. PloS One. 15, e0241871. doi: 10.1371/journal.pone.0241871
Shilling, E., N., Combs, I., R. & Voss, J., D. (2021) Assessing the effectiveness of two intervention methods for stony coral tissue loss disease on Montastraea cavernosa. Sci. Rep. 11, 1-11. doi: 10.1038/s41598-021-86926-4
Slavov, C., Schrameyer, V., Reus, M., Ralph, P., J., Hill, R., Büchel, C., Larkum, A., W., D. & Holzwarth, A., R. (2016) “Super-quenching” state protects Symbiodinium from thermal stress — Implications for coral bleaching. Biochimica et Biophysica Acta. 1857, 840-847.
Smith, H., A., Fulton, S., E., McLeod. I., M. & Bourne, D., G. (2023) Sea-weeding: Manual removal of macroalgae facilitates rapid coral recovery. Journal of Applied Ecology. 00, 1-13. DOI: 10.1111/1365-2664.14502
Sodhi, K., K., Kumar, M. & Singh, D., K. (2021) Insight into the amoxicillin resistance, ecotoxicity, and remediation strategies. Journal of Water Process Engineering. 39, 101858. https://doi.org/10.1016/j.jwpe.2020.101858Get rights and content
Soffer, N., Brandt, M., Correa, A., M., S., Smith, T., B. & Thurber, R., V. (2013) Potential role of viruses in white plague coral disease. The Isme Journal. 8, 271-283.
Starcevic, A., Dunlap, W., C., Cullum, J., Shick, J., M., Hranueli, D. & Long, P., F. (2010) Gene expression in the scleractinian acropora microphthalma exposed to high solar irradiance reveals elements of photoprotection and coral bleaching. PLoS ONE. 12. 5(11), 13975.
Studivan, M., S., Baptist, M., Molina, V., Riley, S., First, M., Soderberg, N. et al. (2022a) Transmission of stony coral tissue loss disease (SCTLD) in simulated ballast water confirms the potential for ship-born spread. Sci. Rep. 12, 19248. doi: 10.1038/ s41598022-21868-z
Studivan, M., S., Eckert, R., J., Shilling, E., Soderberg, N., Enochs, I., C. & Voss, J., D. (2023) Stony coral tissue loss disease intervention with amoxicillin leads to a reversal of disease-modulated gene expression pathways. Mol. Ecol 32 (19), 5294-5413. doi: 10.1111/ mec.17110
Studivan, M., S., Rossin, A., M., Rubin, E., Soderberg, N., Holstein, D., M. & Enochs, I., C. (2022b) Reef Sediments Can Act As a stony coral tissue loss disease vector. Frontiers in Marine Science.
Sunagawa, S., DeSantis, T., Z., Piceno, Y., M., Brodie, E., L., DeSalvo, M., K., Voolstra, C., R., Weil, E., Andersen, G., L. & Medina, M. (2009) Bacterial diversity and White Plague Disease-associated community changes in the Caribbean coral Montastraea faveolata. The Isme Journal. 3, 512-521.
Sutherland, K., P. & Ritchie K., B. (2004) White Pox Disease of the Caribbean Elkhorn Coral, Acropora palmata. Coral Health and Disease. Rosenberg, E. & Loya, Y. (eds.). Springer, Berlin, Heidelberg. pp 289-300.
Sutherland, K., P., Berry, B., Park, A., Kemp, D., W., Kemp, K., M., Lipp, E., K. & Porter, J., W. (2016) Shifting white pox aetiologies affecting Acropora palmata in the Florida Keys. Philosophical Transactions of the Royal Society B: Biological Sciences. 371(1689),.
Sutherland, K., P., Shaban, S., Joyner, J., L., Porter, J., W. & Lipp, E., K. (2011) Human Pathogen Shown to Cause Disease in the Threatened Eklhorn Coral Acropora palmata. PLoS ONE. 6(8), 23468.
Tait, K., Hutchison, Z., Thompson, F., L. & Munn, C., B. (2010) Quorum sensing signal production and inhibition by coral‐associated vibrios. Environmental Microbiology Reports. 2, 145-150.
Takada, Y., Ye, X. & Simon, S. (2007) The integrins. Genome Biol. 8, 215. doi: 10.1186/gb-2007-8-5-215
Tchernov, D., Gorbunov, M., Y., de Vargas, C., Narayan Yadav, S., Milligan, A., J., Häggblom, M. et al. (2004) Membrane lipids of symbiotic algae are diagnostic of sensitivity to thermal bleaching in corals. Proc. Natl. Acad. Sci. 101, 13531-13535. doi: 10.1073/pnas.0402907101
Teplitski, M. & Ritchie, K. (2009) How feasible is the biological control of coral diseases? Trends in Ecology & Evolution. 24(7), 387-385.
Thome, P., E., Rivera-Ortega, J., Rodrıguez-Villalobos, J., C., Cerqueda-Garcıa, D., Guzman Urieta, E., O., Garcıa-Maldonado, J., Q. et al. (2021) Local dynamics of a white syndrome outbreak and changes in the microbial community associated with colonies of the scleractinian brain coral Pseudodiploria strigosa. PeerJ. 9, e10695. doi: 10.7717/peerj.10695
Thornhill, D., J., Kemp, D., W., Bruns, B., U., Fitt, W., K. & Schmidt, G., W. (2008) Correspondence Between Cold Tolerance and Temperate Biogeography. A Western Atlantic Symbiodinium (Dinophyta) Lineage 1. J. Phycology. 44, 1126-1135. doi: 10.1111/j.1529-8817.2008.00567.x
Titus, K., O’Connell, L., Matthee, K. & Childress, M. (2022) The influence of foureye butterflyfish (Chaetodon capistratus) and symbiodiniaceae on the transmission of stony coral tissue loss disease. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.800423
Toledo-Hernandez, C. & Ruiz-Diaz, C., P. (2014) The immune responses of the coral. Inverteb. Surv. J. 11, 319-328.
Toledo-Hernandez, C., Ruiz-Diaz, C., P., Ramirez-Lugo, J., S., Torres-Diaz, M., Santiago-Pagan, L., Bruno-Chardon, A. & Diaz-Vazquez, L., M. (2023) Uncovering the link between environmental factors and coral immunity: A study of fluorescent protein expression and phenoloxidase activity in Acropora cervicornis. Forntiers in Marine Scinence. https://dio.org/10.3389/fmars.2023.1133486
Toledo-Hernández, C., Zuluaga-Montero, A., Bones-González, A., Rodríguez, J., A., Sabat, A., M. & Bayman, P. (2008) Coral Reefs. 27, 707.
Tonk, L., Sampayo, E., LaJeunesse, T., Schrameyer, V., Hoegh‐Guldberg, O. & Mock, T. (2014) Symbiodinium (Dinophyceae) diversity in reef‐invertebrates along an offshore to inshore reef gradient near Lizard Island, Great Barrier Reef. Journal of Phycology. 50(3), 552-563.
Traylor-Knowles, N., Connelly, M., T., Young, B., D., Eaton, K., Muller, E., M., Paul, V., J., et al. (2021) Gene Expression Response to Stony Coral Tissue Loss Disease Transmission in M. cavernosa and O. faveolata From Florida. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.681563
Truc, M., Rivera, A., Ochoa, G., M., Dueñas, D., Guifarro, Z., Brady, G. et al. (2023) Evaluating the spread of stony coral tissue loss disease in the Bay Islands, Honduras. Front. Mar. Sci. 10. doi: 10.3389/fmars.2023.1197318
Ushijima, B., Gunasekera, S., P., Meyer, J., L., Tittl, J., Pitts, K., A., Thompson, S. et al. (2023) Chemical and genomic characterization of a potential probiotic treatment for stony coral tissue loss disease. Commun. Biol. 6, 248. doi: 10.1038/s42003-023-04590-y
Ushijima, B., Meyer, J., L., Thompson, S., Pitts, K., Marusich, M., F., Tittl, J. et al. (2020) Disease diagnostics and potential coinfections by vibrio coralliilyticus during an ongoing coral disease outbreak in Florida. Front. Microbiol. 11. doi: 10.3389/fmicb.2020.569354
Ushijima, B., Smith, A., Aeby, G., S. & Callahan, S., M. (2012) Vibrio owensii Induces the Tissue Loss Disease Montipora White Syndrome in the Hawaiian Reef Coral Montipora capitata. PLoS ONE. 7(10), 46717.
van de Water, J., A., J., M., Ainsworth, T., D., Leggat, W., Bourne, D., G., Willis, B., L. & van Oppen, M., J., H. (2015) The coral immune response facilitates protection against microbes during tissue regeneration. Molecular Ecology. 24, 3390-3404. https://doi.org/10.1111/mec.13257
van de Water, J., A., J., M., Chaib De Mares, M., Dixon, G., B., Raina, J., B., Willis, B., L., Bourne, D., G., van Oppen, M., J., H. (2018) Antimicrobial and stress responses to increased temperature and bacterial pathogen challenge in the holobiont of a reef-building coral. Mol. Ecol. 27, 1065-1080. https://doi.org/10.1111/mec.14489
van de Water, J., A., J., M., Lamb, J., B., Heron, S., F., van Oppen, M., J., H. & Will, B., L. (2016) Temporal patterns in innate immunity parameters in reef-building corals and linkages with local climatic conditions. Ecosphere 7 (11), e01505. https://doi.org/10.1002/esc2.1505
van Oppen, M., J. & Burghardt, I. (2009) Zooxanthellae (Symbiodinium, Dinophyceae) symbioses on coral reefs. Microbiol. Aust. 30, 67. doi: 10.1071/MA09067
Veglia, A., J., Beavers, K., Van Buren, E., W., Meiling, S., S., Muller, E., M., Smith, T., B. et al. (2022) Alphaflexivirus genomes in stony coral tissue loss disease-affected, disease exposed, and disease-unexposed coral colonies in the U.S. Virgin islands. Microbiol. Resource Announcements. 11, e01199–e01121. doi: 10.1128/mra.01199-21
Veron, J., E., N. (1995) Corals in space and time. Veron, J., E., N. (ed.). UNSW Press, Sydney.
Vidal-Dupiol, J., Dheilly, N., M., Rondon, R., Grunau, C., Cosseau, C., Smith, K., M., Freitag, M., Adjeroud, M. & Mitta, G. (2014) Thermal stress triggers broad Pocillopora damicornis transcriptomic remodeling, while Vibrio coralliilyticus infection induces a more targeted immuno-suppression response. PLoS One. 9, e107672. https://doi.org/10.1371/journal.561 pone.0107672
Vidal-Dupiol, J., Ladrière, O., Meistertzheim, A., L., Fouré, L., Adjeroud, M. & Mitta, G. (2011) Physiological responses of the scleractinian coral Pocillopora damicornis to bacterial stress from Vibrio coralliilyticus. The Journal of experimental biology. 214(9), 1533-1545.
Voolstra, C., R. & Ziegler, M. (2020) Adapting with Microbial Help: Microbiome Flexibility Facilitates Rapid Responses to Environmental Change. BioEssays : news and reviews in molecular, cellular and developmental biology. 42(7), e2000004.
Walker, B., K., Turner, N., R., Noren, H., K., G., Buckley, S., F. & Pitts, K., A. (2021) Optimizing Stony Coral Tissue Loss Disease ( SCTLD ) Intervention Treatments on Montastraea cavernosa in an Endemic Zone. 8, 1-11. doi: 10.3389/ fmars.2021.666224
Wallace, C., C. & Rosen, B., R. (2006) Diverse staghorn corals (Acropora) in high-latitude Eocene assemblages: Implications for the evolution of modern diversity patterns of reef corals. Proceedings of the Royal Society B: Biological Sciences. 273, 975-982.
Wegley, L., Edwards, R., Rodriguez-Brito, B., Liu,H. & Rohwer, F. (2007) Metagenomic analysis of the microbial community associated with the coral Porites astreoides. Environ. Microbiol. 9, 2707-2719. doi: 10.1111/j.1462 2920.2007.01383.x
Weil, E. (20040 Coral Reef Diseases in the Wider Caribbean. Coral Health and Disease Rosenberg, E. & Loya, Y. (eds.). Springer. pp 35-68. https://doi.org/10.1007/978-3-662-06414-6_2
Weinberg, E. (2018) Scientists work together to solve a coral disease mystery in Florida Keys. National Marine Sanctuary. https://sanctuaries.noaa.gov/news/aug18/coral-disease-mystery-florida-keys.html
Weisburg, W., G., Barns, S., M., Pelletier, D., A. & Lane, D., J. (1991) 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriology 173, 697-703. doi: 10.1128/ jb.173.2.697-703.1991
Williams, S., D., Walter, C., S. & Muller, E., M. (2021a) Fine scale temporal and spatial dynamics of the stony coral tissue loss disease outbreak within the lower Florida keys. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.631776
Williams, S., M., Garcıa-Sais, J. & Sabater-Clavell, J. (2021b) Prevalence of Stony Coral Tissue Loss Disease at El Seco, a Mesophotic Reef System off Vieques Island, Puerto Rico. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.668669
Williamson, O., M., Dennison, C., E., O’Neil, K. L. & Baker, A. C. (2022) Susceptibility of Caribbean brain coral recruits to stony coral tissue loss disease (SCTLD). Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.821165
Wirth, J., S., Wang, T., Huang, Q., White, R., H. & Whitman, W., B. (2020) Dimethylsulfoniopropionate Sulfur and Methyl Carbon Assimilation in Ruegeria Species. mBio. 11(2), e00329-00420.
Woese, C., R. & Fox, G., E. (1977) Phylogenetic structure of the prokaryotic domain: the primary kingdoms. Proc. Natl. Acad. Sci. U.S.A. 74, 5088-5090. doi: 10.1073/pnas.74.11.5088
Woese, C., R. (1987) Bacterial evolution. Microbiological Rev. 51, 221-271. doi: 10.1128/mr.51.2.221-271.1987
Work, T., M., Weatherby, T., M., Landsberg, J., H., Kiryu, Y., Cook, S., M. & Peters, E., C. (2021) Viral-like particles are associated with endosymbiont pathology in Florida corals affected by stony coral tissue loss disease. In: Frontiers in Marine Science. 8, 1-18.
Xu, M., Cheng, K., Xiao, B., Tong, M., Cai, Z., Jong, M-C., Chen, G. & Zhou, J. (2023) Bacterial Communities Vary from Different Scleractinian Coral Species and between Bleached and Non-Bleached Corals. Microbiology Spe ctrum. 11(3),.
Yu, J., Liao, T., Chen, C., Liao, T., Fang, L. & Tsai, W. (2000) Do color patterns of Pocillopora damicornis reflect Zooxanthellae diversity? Coral Reefs. 19(1), 98-99.
Zhao, M., Zhang, H., Zhonf, Y., Xu, X., Yan, H., Li, G. & Yan, W. (2021) Microstructural characteristics of the stony coral genus Acropora useful to coral reef paleoecology and modern conservation. Ecology & Evolution. 11, 3093-3109. http://www.dio.org/10.1002/ece3.7247
Zhou, Z., Yu, X., Tang, J., Zhu, Y., Chen, G., Guo, L. et al. (2017) Dual recognition activity of a rhamnose-binding lectin to pathogenic bacteria and zooxanthellae in stony coral Pocillopora damicornis. Dev. Comp. Immunol. 70, 88-93. doi: 10.1016/ j.dci.2017.01.009
Zoccola, D., Ounais, N., Barthelemy, D., Calcagno, R., Gaill, F., Henard, S., Hoegh-Guldberg, O., Janse, M., Jaubert, J., Putnam, H., Salvat, B., Voolstra, C. & Allemand, D. (2020) The World Coral Conservatory (WCC): A Noah’s ark for corals to support survival of reef ecosystems. PLoS Biology. 18(9),
To learn more, visit https://reefranch.co.uk/
![]()

0 Comments