

Favites (complanata?). Credit: unknown
Merulinidae: Favites
With twenty species, this is the second most speciose genus of merulinid coral. As discussed above, Favites shares most of its morphological characters with Dipsastraea, differing most visibly in the nature of the corallite arrangement (typically ceriod vs plocoid), though this too is variable and often unreliable. A good example is the placement of the former “Favia rotundata” within Favites based on molecular study. Confusing matters is the placement of several Indo-Pacific species formerly included in Montastraea, which stretches the definition of the genus beyond a tidy, easy-to-understand description. Also note that there are microstructural differences between Favites and Dipsatraea related to skeletal calcification that can provide a more authoritative means to identify these corals, but this is beyond the means of any sane aquarist.

F. valenciennesi, note the extracalicular budding and uneven septa. Credit Franchi Cichlids
Favites is also strikingly similar to several other genera that possess cerioid polyps: Goniastrea, Paragoniastrea, Platygyra and Coelastrea. As a general rule of thumb: Goniastrea has less calice relief and usually has prominent septal lobes (vs weaker paliform lobes in Favites); Platygyra has more irregularly exsert septa, giving it a ragged look; Coelastrea has taller thecal walls with septa that give it an evenly-ribbed appearance.

Favites pentagona, the “war coral”. Credit: Exotic Reef Creations
Favites species can be combined into a few informal groups based on polyp size and shape. Three species formerly placed in Montastraea (colemani, magnistellata, valenciennesi) are identifiable by their plocoid polyps and extracalicular budding. They are easily confused with Dipsastraea, but the differences in budding should easily separate the two. While generally uncommon (or overlooked) in aquarium exports, all three have wide geographic ranges and are by no means rare in the wild.
Another distinctive group is formed by three species with small (< 6mm) angular polyps. The most common species is F. pentagona (stylifera and micropentagona are less likely to be seen). Known commonly as the “war coral”, this species appears as thin, laminar colonies with variable amounts of vertical branching towards the center. The branching is generally an indication of the colony originating from an area of the reef with low water movement. Coloration is usually a brick red with variable amounts of marbling and bright green oral disks. Note that there is molecular evidence to indicate F. pentagona is distinct from other Favites studied to date, and it may well be elevated to genus rank with further study.
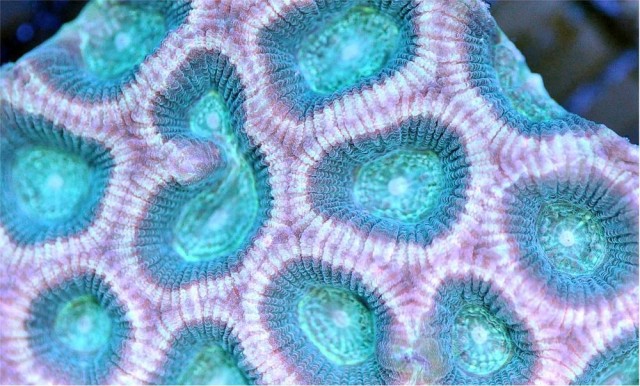
Favites flexuosa?, the “reverse prism favia”. Credit: Aqua San Diego
The final informal group is composed of those species not already discussed. These are corals possessing cerioid polyps of medium to large size. Some species (complanata and abdita) are common exports, while many of the others are rarely, if ever, seen. Species level identifications are challenging, and confusion with monocentric species of Goniastrea, Platygyra and Coelastrea is an easy mistake to make.

Goniastrea pectinata, note the septal lobes. Credit: Guam Marine Lab
Merulinidae: Goniastrea
This genus has been a consistent source of confusion for taxonomists and aquarists alike. To give an idea of this uncertainty, species that have at one time been classified within Goniastrea are now placed in seven different genera. Aquarium references typically identify this group as resembling Favites, but with more prominent paliform lobes. This is a gross oversimplification that overlooks the significant variability of this feature in both groups, as well as the fundamental differences (i.e. septal vs paliform lobes) between these genera.

G. favulus, with a strongly meandroid skeleton. Credit: Ryan McMinds
G. retiformis and minuta have small (3-5mm), monocentric polyps, which gives the thecal ridges of the colony a net-like look. They somewhat resemble F. pentagona, but differ in coloration and colony shape (massive instead of laminar/branched). G. edwardsi is another similar species common on reefs. Polyps average slightly larger (2-7mm) and tend to have thicker walls. G. pectinata and favulus vary from cerioid to submeandroid, with particularly meandroid specimens of favulus strongly resembling P. australensis. (These can be told apart by the thicker ridges and narrower valleys in the common aquarium species P. australensis.) G. pectinata is reported to be the most common species in the wild, while favulus is mostly reported from intertidal habitats.

Goniastrea (retiformis?), note the septal lobes (green). Credit: House of Fins
Two aberrant species round out this group. G. ramosa is a rare species on reefs that grows as vertical branches, strongly reminiscent of Australogyra zelli. G. stelligera is a surprising addition to this genus, as it strongly resembles Dipsastraea in possessing plocoid polyps with an expansive coenosteum. The differences that align this species with Goniastrea are primarily microstructural, though it bears mention that none of the species in Dipsatraea have polyps quite so small. G. stelligera is reported to be a common species on reefs, but due to its large columnar growth form and pale tan coloration, it is unlikely this species is ever exported.
Goniastrea is still a difficult group to identify. It’s uncertain how often they appear for sale, as more often than not they are sold unidentified or misidentified. While their general structure is not dissimilar to Favites, the cerioid Goniastraea are typically much smaller in polyp diameter, with less calice relief and more prominent septal lobes. Certain Platygyra are likewise similar, but can typically be told apart due to their more variably exsert septa. Coelastrea are again similar, but have larger polyps and far greater calice relief than in Goniastrea.

Coelastrea aspera, note the steep, thin walls. Credit: Pacific Aqua Farms
Merulinidae: Coelastrea
Two species are definitively known for this group, aspera and palauensis, and a third species, tenuis, is known only from the holotype specimen that established the genus in 1866. This specimen is nearly identical to aspera and raises serious questions about its existence as anything more than a taxonomic artifact.

C. aspera, typical honeycombed appearance, exposed at low tide. Credit: Melissa Copnell
Authors have vacillated on whether or not these species should be lumped into Goniastrea, with most recent sources choosing to combine the groups. But molecular study indicates this is a distinct lineage, most closely related to two genera with a very different appearance, Dipsatraea and Trachyphyllia. The differences used to diagnose between the superficially similar Coelastrea and Goniastrea involve microstructural nuances that are relevant to only the most pedantic of coral taxonomists. A helpful rule of thumb for aquarists is that Coelastrea typically have larger polyps (7-10mm) and taller thecae than Goniastrea.
C. aspera is a common species associated with intertidal parts of the reefs, meaning this species is regularly exposed during low tides. The common name “honeycomb coral” has been applied to this distinctive species, as the rounded, boulder-shaped colonies possess tall, thin thecal walls that give the polyps a “deep and cellular” appearance. Coloration of aquarium specimens attributable to this species have thin, brown walls and bright green oral disks.

C. palauensis, a common green form. Credit: Matt @ Dallas/Fort Worth MAS
Its congener, C. palauensis, is quite similar in appearance but has noticeably thicker walls. Colonies in the wild are reported to grow to a maximum of around 20cm across, with a variably hemispherical shape. There seem to be at least five color morphs of this species in the aquarium trade: 1) solid neon green 2) solid neon/dull green with contrasting teal oral disks 3) solidly mauve/purple 4) maroon with whitish oral disks 5) marbled green and brown.
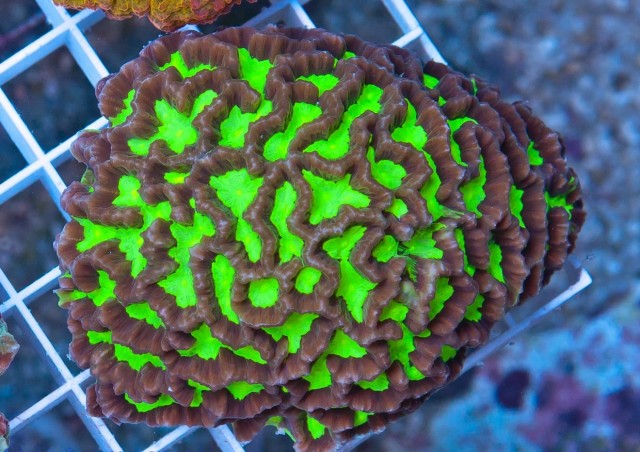
Platygyra (acuta?), note the uneven septa. Credit: Extreme Corals
Merulinidae: Platygyra
This highly variable genus of 11 species is difficult to define but is often easy to identify. The polyp shape can vary between and within species, from cerioid to submeandroid to extensively meandroid “maze corals”. Identifying to species level is wrought with difficulties given the potential variation within a species, but the group can usually be diagnosed by the rather uneven appearance of the septa, which often alternate in relative height (low, high, low, high…).

P. daedalea, note the prominent septa and wide valleys. Credit: Ike Eigenbrode/Ecoxotic
P. daedalea is the quintessential maze coral, with the polyps arranged in short, highly meandering valleys. The species is named after the architect of the famous labyrinth of Ancient Greek mythology, Daedalus—one of the more evocative etymological allusions in coral nomenclature. P. lamellina is a similar species that differs in its thicker walls, longer valleys and unusually even septa (for the genus). Both species resemble Paragoniastrea australensis, but it differs in possessing prominent paliform lobes and discrete columellae. To best observe the presence of paliform lobes try disturbing the colony into deflating its tissue, as fully inflated specimens can easily hide this feature.

Platygyra (verweyi?) Credit: RedClay/reefcentral
The remaining species are a heterogeneous mix showing either primarily cerioid or submeandroid polyps, though it seems that these disparate growth forms can appear variably in most species under the right environmental conditions. P. pini and verweyi are examples of primarily cerioid species that appear sporadically in aquarium stores; specimens which I have encountered were a deep green and produced extremely long (~20cm) sweeper tentacles. The thecae are thinner and more truncated in verweyi and more rounded in pini. Identification to species in this group is extraordinarily challenging, and likely beyond the capabilities of all but the most dedicated of coral researcher. The literature is filled with synonyms that attests to that.
Not many corals will be confused with Platygyra. Cerioid specimens do bear some resemblance to Favites, but the polyps of Platygyra are generally far smaller (~5mm) and with irregularly shaped septa. Goniastrea is also superficially similar, but the polyps of Goniastrea differ in showing less calice relief (< 3mm), possessing large septal lobes and having more evenly shaped septa.
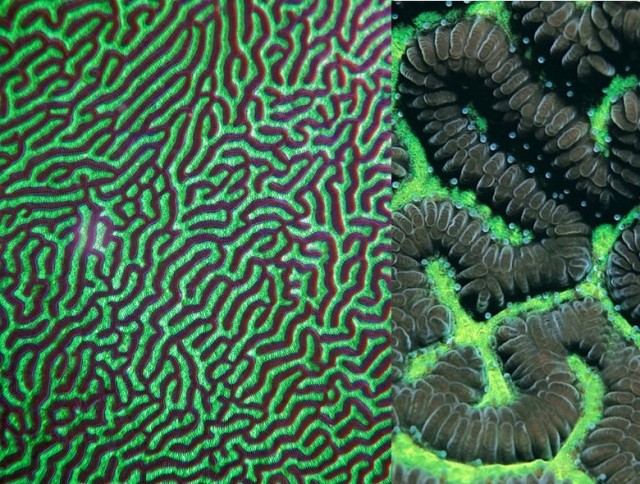
L. phrygia, note the even, medially-abutting, zipper-like septa. Credit: Simon Grove
Merulinidae: Leptoria
Two species of “maze” or “worm” brain corals compose this genus. The common species L. phrygia forms large, massive colonies with narrow, meandering valleys. These valleys are often far more sinuous than similar meandroid species, and the valleys are particularly small (3-4mm), often narrower in width than the ridges. The septa are evenly shaped and abut medially atop the ridges, giving them a zipper-like look. This species can easily be confused with Platygyra sinensis, as both have narrow and highly meandering valleys; the septal differences will help differentiate the two. Coloration of exported specimens is usually a bright green valley with brown ridges, though some specimens may be a drab mauve-brown.
Beneath the tissue, L. phrygia has an unusually shaped columella, forming short vertical sheets (i.e. laminar) as opposed to the typical spongy mass of nearly all merulinds. While common on reefs, this species doesn’t appear very often for sale, presumably due to the need to harvest suitably small colonies for export. This is in stark contrast to the numerous retailers that offer misidentified specimens of Platygyra or Paragoniastrea under the name Leptoria. Nearly every online image I’ve seen from the aquarium trade purporting to be Leptoria is incorrectly identified. Caveat emptor!

Platygyra sinensis, note the uneven, mostly non-abutting septa. Credit: rustl3r
The second species in the genus is L. irregularis. It bears a resemblance to phrygia, but differs in possessing a spongy columella and more irregularly shaped septa. It seems this species is significantly rarer than phrygia, and the coloration is only reported as a pale brown. Presumably this species doesn’t appear in the aquarium hobby. It could easily be confused with the more meandering species of Platygyra, though the relative size of the ridges and valleys should tell it apart. The habitat for both Leptoria species is primarily the upper reef slope, where water flow is typically quite high. It’s a good idea to avoid placing them in low flow regions of an aquarium.
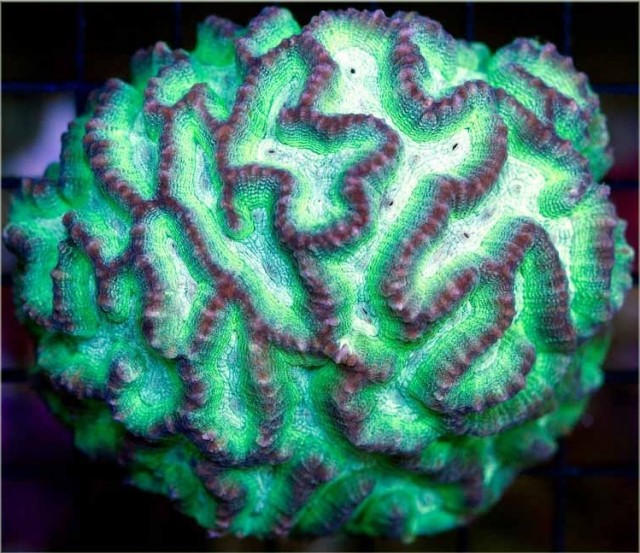
Oulophyllia, note the large, exsert septa and wide, sloping valleys. Credit: Aqua San Diego
Merulinidae: Oulophyllia
Three species comprise this group. O. levis and crispa form massive, hemispherical colonies with large (10-20mm) polyps arranged into short meandering valleys. These valleys are considerably larger than the other meandering merulinids, and are further identifiable by their strongly sloping, V-shaped walls. There are relatively fewer septa than in related genera, which results in the ridges appearing prominently bumpy. Coloration ranges from a bluish-grey to green, and they are an uncommon find, both on the reef and in aquariums.

O. bennettae. Credit: unknown
O. bennettae differs from its congeners in being primarily monocentric, but is otherwise quite similar. It could potentially be mistaken for a large-polyped species of Favites, but the widely-spaced septa and bumpy appearance, as well as the bluish-grey color, help to diagnose it. Again, this is a rarely seen species amongst aquarists, but it does occasionally get exported.
An unusual behavior found in this genus is the ability to release asexually formed buds, in a similar manner as is seen in Pocillopora and Tubastraea. This has given rise to the amusing common name “ejector seat coral” in some publications.

P. australensis, typical coloration of aquarium specimens. Credit: Golden Ina
Merulinidae: Paragoniastraea
Three species form this recently described genus. These have traditionally been placed in either Goniastrea or Favites, and it should come as little surprise that they are challenging to tell apart. The most distinctive species, and the one most likely to appear in aquariums, is P. australensis. Aquarium specimens normally have long meandering valleys of variable width, and ridges that range from low and angular to high and steep. Unlike similar meandering species (e.g. Leptoria phyrgia, Platygyra lamellina), this species often has short meanders, or even an occasional monocentric corallite.

P. australensis, another common color pattern; specimen submeandroid. Credit: Corals Galore
The colony shape of australensis is normally either submassive or encrusting and platelike. Exported specimens are normally thin, irregular pieces broken from larger colonies, but small, submassive colonies do appear on rare occasions. Coloration comes in three common flavors: 1) brown ridges and bright whitish valleys 2) solid dull green 3) neon green septa on dark ridges, with either green or dark oral valleys.

P. russelli, note the jagged septa and tricoloration. Credit: Pacific Aqua Farms
P. russelli is a challenging species to identify, as it bears a resemblance to both Favites (where it was formerly placed) and Platygyra. Its cerioid polyps with irregularly shaped septa are indeed Platygyra-like, but its strong paliform lobes tell these two apart. The combination of uneven septa and prominent paliform lobes is also useful for delimiting russelli from Favites. Confusingly, this species can also appear subplocoid in turbid environments, which could lead to confusion with Dipsatraea. Veron reports this species as common on the Great Barrier Reef, and it has a wide range throughout the Indo-Pacific. Aquarium specimens can be mostly green or have contrasting maroon oral disks (as above).
The last species to discuss is P. deformis, a rarely encountered species in the wild with a wide geographic range. This species is reminiscent of Goniastrea, but has more calice relief (> 3mm vs < 3 mm). It is reported primarily from Southern Japan and the Western Indian Ocean, and colonies are a drab brown.

P. peresi. Credit: Dr. Charlie Veron
Merulinidae: Paramontastraea
The three species in this newly formed genus (salebrosa, peresi, serageldini) are somewhat tentatively grouped and have traditionally been placed in either Goniastrea or Montastraea. More than any other group discussed here, this genus may not yet be taxonomically stable.
P. salebrosa and serageldini are similar, both possessing small, extracalicularly-budded polyps. They also share the unusual condition of a spinose coenosteum, a trait shared with Cyphastraea and the Atlantic Orbicella. P. salebrosa is found in the Pacific, but its drab brown coloration makes it unlikely to ever be exported. P. serageldini is restricted to the West Indian Ocean, and thus unavailable. The real oddball in this group is P. peresi, which bears little resemblance to its congeners, but yet it groups with them (tentatively) in molecular studies. P. peresi has strongly cerioid polyps and prominent paliform lobes (hence its former inclusion in Goniastrea). Budding is variably intra- or extracalicular. Its range is restricted to the Indian Ocean, where it is reported to be a common species.
















0 Comments