

Iodine is a component of seawater that has long captured the interest of marine aquarists. There are many commercial iodine supplements intended for aquaria, and a lot of commentary about what it is supposed to be good for (name your critter: shrimp, Xenia, mushrooms, soft corals in general, macroalgae, etc.). Others express the opinion that supplementation is unnecessary and possibly dangerous. Most aquarists aren’t sure whether supplementation helps anything, and these folks are about equally split between those who dose iodine “just in case” and those who do not.
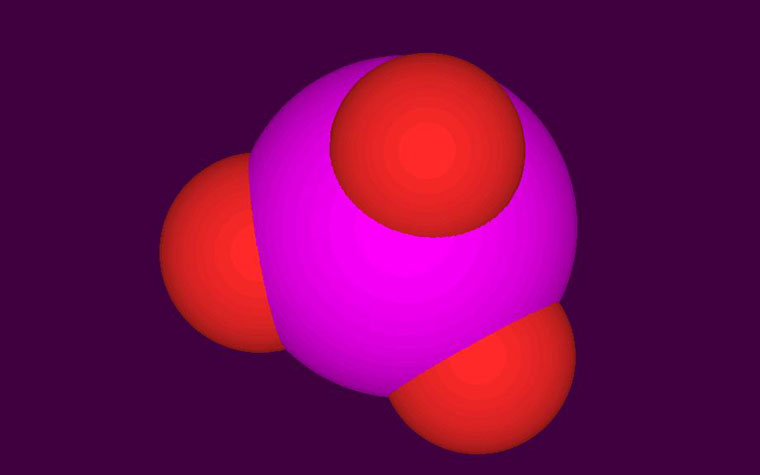
Figure 1. A space filling model of an iodiate ion (IO3-). Note how large the central iodine atom is compared to the three oxygen atoms.
This article is the first of several that attempt to bring some small amount of clarification to the world of iodine in aquaria. Together these articles will expand on the article that Craig Bingman published six years ago on iodine in seawater and aquaria.1 This first article will cover what is known about iodine in the oceans, including what forms it takes and how toxic these forms are, what organisms use it, how they obtain it, and what they use it for. It will also detail some issues around iodine measurement and what natural sources of iodine are significant to aquaria.
Future articles will attempt to show how rapidly iodide and iodate are depleted from aquaria and discuss where it might be going. Whether to dose an iodine supplement or not, and if so, what to dose and how much, are questions that are much more complicated than they might seem (and much more complicated than I believed in the past). There are already big natural inputs of iodine to many marine aquaria, as well as significant exports. With all of the different forms of iodine coming and going at different rates, with the different forms rapidly interconverting, with the different forms having different availability to organisms that take them up, and with different test methods detecting different forms differently, deciding what and how much to dose, if at all, is not a simple case of trying to match natural seawater (a worthy goal, but in this case, perhaps impossible for most aquarists to attain).
While the scientific literature is loaded with articles that describe how and why many marine organisms use iodine, it is silent, not surprisingly, on whether supplementation of iodine in normal marine aquaria results in anything that the aquarist would call positive. As will be shown below, many macroalgae grow more rapidly in the presence of iodine, and some require it. In an effort to test some likely hypotheses of how iodine supplementation may be beneficial to real aquaria, the second article will report on some studies of how iodine supplementation impacts the growth of two macroalgae often selected for rapid growth in refugia: Caulerpa racemosa and Chaetomorpha sp.
In this article I won’t make a specific recommendation concerning dosing, since the results of the various studies to be reported in future articles (some of which are already completed, but some not) will impact that recommendation. I do suggest in this article, however, that there are some practices that aquarists employ involving iodine that I think have the potential to lead to problems, and that therefore should be avoided.
Iodine in the Ocean
Iodine in the ocean takes a wide variety of forms, both organic and inorganic, and the iodine cycles between these various compounds are very complex and are still an active area of research. The nature of inorganic iodine in the oceans has been generally known for decades. The two predominate forms are iodate (IO3–, with the central iodine and three attached oxygen atoms; Figure 1) and iodide (I–). As a curiosity, note the huge size of the iodine atom compared to the oxygen atoms in iodate in Figure 1. Together these two iodine species usually add up to about 0.06 ppm total iodine (~0.5 mM), but the reported values vary over about a factor of 2. In surface seawater, iodate usually is the dominant form with typical iodate values in the 0.04 to 0.06 ppm iodine (0.3 – 0.5 mM).2,3 Likewise, iodide is usually present at lower concentrations, typically 0.01 to 0.02 ppm iodine (0.07 – 0.18 mM).2,3
Organic forms of iodine are any in which the iodine atom is covalently attached to a carbon atom, such as methyl iodide, CH3I. The concentrations of the organic forms (of which there are many different molecules) are only now becoming recognized by oceanographers. In some coastal areas, organic forms can comprise up to 40% of the total iodine, and many previous reports of organoiodine compounds being negligible may be incorrect.4 Later in this article, some of the organoiodine species found in the ocean are discussed in the context of those organisms that produce them.
All of these various forms can be interconverted in the oceans. Phytoplankton, for example, take up iodate and convert it into iodide, which is mostly, but not completely, released.5 One research group has suggested that iodate, looking chemically like nitrate, is taken up by the same pathways, and is internally converted to iodide before being released.6 This process is fast enough that in one local studied, the phytoplankton can convert all of the iodate present to iodide in a month. Iodate is also converted to iodide by bacteria in low oxygen environments of the oceans.7
Marine algae can also take up iodide directly, and apparently do so preferentially over iodate.8-10 This process may, in fact, be a primary way that iodide is depleted from aquaria, but that’s getting ahead of things.
There are also abiotic (nonbiological) transformations taking place in the oceans, with iodide being potentially oxidized to iodate. These abiotic processes are probably not the controlling factors of iodine speciation in the oceans, however, with biological processes predominating.11 In marine aquaria that employ strong oxidants such as ozone, or possibly even UV sterilizers that can promote oxidation, these abiotic factors may predominate.
Toxicity and other properties of various forms of iodine
The various forms of iodine have very different chemical properties, and they should not be confused with each other. In addition to the three forms mentioned above, aquarists also frequently encounter molecular iodine (I2). Lugols’ solution, for example, contains a mixture of I2 and I–. It is the I2 form in particular that is the basis for the widespread belief that iodine is “toxic.” The I2 form, and that form in combination with other forms, it is a potent antibacterial agent that has long been used for disinfection. The naturally occurring inorganic forms (iodide and iodate) have little in the way of antimicrobial activity.12 Even a 30,000 ppm solution of iodide (240 mM or half a million times more concentrated than in normal seawater) is only weakly antibacterial13 In mixtures containing I2 and other forms, it is the amount of free I2 that is important for antimicrobial activity.14
In terms of toxicity to higher organisms, the concerns vary considerably from organism to organism. Still, these general trends seem to hold. Rainbow trout, for example, are quite sensitive to I2, with the LC50 (the concentration where half of them die) below 1 ppm iodine. They are not very sensitive to I– or IO3–, with the LC50 for these species being greater than 200 and 850 ppm respectively.15,16 Daphnia magna were equally sensitive to I2 (LC50 less than 0.2 ppm) and I– (LC50 less than 0.2 ppm), but were less sensitive to IO3– (LC50 above 10 ppm ) .15
Unfortunately, there is not very much toxicity data available for any of these iodine species on marine organisms. Aquarists are left not knowing exactly how high the iodide and iodate levels can get before becoming problematic. Phytoplankton growth is apparently not inhibited at iodate levels up to 1.3 ppm iodine (iodide, which they made from the iodate was also presumably high).17 These authors concluded “there is little interaction between iodine processing and the metabolic activity of cell growth.” Several species of phytoplankton were shown to be uninhibited by iodide and iodate at greater than 12 ppm, though iodide (but not iodate) began to inhibit one species (C. antiqua) at levels below about 0.13 ppm.18
In the subsequent sections, it is made clear that macroalgae often thrive at elevated iodide or iodate levels, sometimes better than at natural levels. Whether this extends to other organisms in marine aquaria is not known.
Who Uses Iodine: Algae
The presence of large quantities of iodine in various species of marine algae has been known for nearly 100 years.19 Some species can contain almost 1% iodine (100,000 ppm) by dry weight. Iodine is concentrated considerably by all macroalgae, including the Rhodophyta (red algae), Phaetophyta (kelp and other brown algae) and the Chlorophyta (green algae). There are, however, great species differences in the amount of iodine contained in the macroalgae tissue. Many of these species are of interest to marine aquarists, either as food for fish, or because they are grown in their tanks.
The amount of iodine present in these macroalgae varies substantially with time of year, water temperature, depth, and other factors.20,21 Some species of macroalgae have a clearly defined requirement for iodine, or else they do not grow. The brown algae Ectocarpus siliculosus, for example, requires at least 0.0015 ppm I– for vegetative growth and 0.0025 ppm I– for normal formation and maturation of Pluriolocular sporangia, though it can survive without growth in iodine deficient solutions. At increased iodine levels, growth increases up to about 0.2 ppm I– , where growth levels off, and stays constant up to at least 20 ppm I–.22 Laminaria saccharina, as another example, shows Meiospore germination increasing with iodine concentration up to about 0.1 ppm, whether it is exposed to iodide or iodate. That
same species is insensitive to iodine concentration after germination.23
The red macroalgae Polysiphonia urceolata apparently shows growth rates that increase linearly with iodine concentration between 0.1 and 1 ppm iodine.24 The same group showed that Nemalion sp. showed no change in growth with iodine concentration, and Goniotrichum elegans was inhibited at iodine concentration above natural levels. The macroalgae Asparagopsis armatahas been shown to grow optimally at iodide or iodate concentrations of 0.6 ppm iodine, and at levels above 1.8 ppm, the growth becomes inhibited.25
Of the macroalgae that marine aquarists most commonly keep, there is no iodine requirement data available (hence the studies to be reported in the second article). Chaetomorpha moniligera has been reported to contain 24 ppm iodine on a dry weight basis,26 Chaetomorpha antennina has been reported to contain 1,100 ppm iodine, and an undetermined Chaetomorpha species contained 360 ppm iodine.27 In a different study, Chaetomorpha antennina and Chaetomorpha linum were shown to contain 144 and 68 ppm iodine, respectively.28
Likewise, Caulerpa racemosa has been reported to contain about 800 ppm iodine by dry weight.20 Caulerpa sertularoide was shown to contain about 310 ppm iodine.27 Caulerpa taxifola has been shown to contain 89 ppm iodine.28 In an aquarium setting, Ron Shimek showed that 3 specimens of Caulerpa sp. contained 440, 843, and 1083 ppm iodine on a dry weight basis.29
What Form Of Iodine Do Macroalgae Take Up?
There have been, in fact, a great many studies on what forms of iodine are taken up. As a consequence of exploding atomic bombs and other nuclear “incidents”, radioactive iodine can be strewn throughout the environment, and a great deal of research has been commissioned by various agencies to study where the radioactive iodine goes. It turns out that macroalgae can take up both iodide and iodate, with a preference for iodide.5, 8-10, 30 The degree of preference seems to vary from species to species, with most able to use both.
What Form Is The Iodine In When Inside Of Macroalgae?
Unfortunately, asking what form iodine takes inside of macroalgae is a much more complicated question than it first sounds. Apparently, macroalgae store iodide in many forms, including both inorganic forms (I– and IO3–, but especially I–) and organic forms, and the amount in these various forms varies from species to species. In one study of many species of macroalgae, the forms soluble in water comprised between 16 and 99% of the total iodine present, and of that water soluble portion, iodide comprised 61-93%, iodate comprised 1.4 to 4.5%, and organic iodine compounds comprised 5.5 to 37%. Presumably, the fraction of the total iodine that is not soluble in water (1-84% of the total iodine) is also comprised of organic iodine compounds.31 Of the common organoiodine compounds found, the iodinated amino acids, monoiodotyrosine and diiodotyrosine, appear prominently.32-34
What benefit is all of this iodine to the macroalgae? That has not been definitively established, but it has been suggested that it is largely an antiherbivore defense. The suggestion is that the algae is either somewhat toxic at these high iodine levels, or simply tastes bad, encouraging the herbivore to move on to something more tasty. The study of trace organoiodine compounds has only recently begun in marine systems, but one recent finding, that an alga in the genus Laurencia contains an iodinated lactone (a metabolite that is likely a poison to herbivores) supports this hypothesis as being at least part of the answer.35
Do Macroalgae Use Iodine In Other Ways?
In addition to storing it inside themselves, algae use iodine in other ways. One prominent way is to produce a variety of organic iodine compounds that are released. These compounds have generated a great deal of study recently because some of them enter the atmosphere and potentially impact global warming and other issues that are generating research dollars. Many of these studies are interested in all types organohalogens (organics containing fluorine, chlorine, bromine, or iodine) that macroalgae produce, but most of those of interest are the 1-4 carbon organics that contain some combination of bromine and iodine.36
Three of the most common iodoorganics of this type are methyl iodide (CH3I), diiodomethane (CH2I2) and iodobutane (C4H9I) .37 It has been hypothesized that these compounds are formed as a byproduct of scavenging hydrogen peroxide inside of cells.37-39 Having such peroxides around is undesirable because they will react with organic materials inside of the cells. In the absence of such scavenging, these peroxides may do substantial damage to the cellular machinery that keeps cells alive, so scavenging peroxide can be an important function. If this is the sole reason for synthesizing these molecules, then the organoiodine compounds do not themselves confer any substantial benefit, but in the absence of making them, the macroalgae may suffer oxidative degradation from peroxide.
In the open ocean, these released small organohalogens are detectable, though typically at fairly low concentrations. In a recent survey of the Atlantic ocean, one research team claimed: “In addition to CH3I, which for a long time was believed to be the only volatile iodinated substance in the marine environment, other iodinated substances like CH2ClI, CH2I2, and CH3CH2CH2I exist in the range of < 0.01-2.2 ng/L in surface water of the Atlantic Ocean.”40 In units that aquarists are familiar with, 2.2 ng/L corresponds to about 0.000002 ppm iodine.
One very recent study, however, has suggested that the monohalomethanes (e.g., CH3I, CH3Br, CH3Cl, but not those organics with multiple halogens or carbon atoms) may simply be a chemical accident41 In their analysis, iodide and bromide may accidentally get into the active site of certain enzymes (various methyltransferases) that are normally serving other functions. As a consequence of this side reaction, these monohalomethanes may be formed. If that hypothesis holds true, then these particular compounds may not serve any useful function at all.
Who Uses Iodine: Gorgonia And Antipatharian Corals (Black Coral)
Another set of creatures of the deep that use iodine are certain gorgonia, such as Plexaura flexuosa.42 They have 3,5-diiodotyrosine in their bodies, to the tune of 0.1 to 2.6% of the total dry weight as iodine. This iodoamino acid is presumably incorporated into proteins in the skeleton (stem), but the benefit is unclear. Again, it may be largely an antipredatory effect that is desired. The iodine incorporation in gorgonia seems to increase with age.43,44 The proteins of many different gorgonia species contain substantial iodine: Eunicella otenocalloides 6.5-8.9% by weight%, Gorgonia verrucosa 4.2-9.0, G. lamarcki 3.3-6.8, G. scirpearia 0.4-0.6, Rhipidigorgia flabellum 0.6-1.1, Euplexora maghrebensis 0.19-0.23, and Plexaura kukenthali 1.9-2.2.44 It has also been demonstrated that at least one gorgonia (E. verrucosa) takes up iodine in the form of inorganic iodine from the water column.45
One study showed that the organoiodine compound thyroxine, and some related compounds, are localized to certain parts of the gorgonia L. virgulata.46 Most interestingly, one of the places it is localized to are scleroblasts (spicule-forming cells) and on the spicules themselves. Further, the addition of thyroxine to these cells impacted the uptake of calcium, and it is suggested that the thyroxine functions in spicule formation.
The antipatharian corals (the black corals) also seem to incorporate a lot of iodine. The basal regions of these corals are especially loaded with iodine, with more than 23% iodine by dry weight recorded in two species.43, 45 Again, the specific purpose is not known.
Who Uses Iodine: Tunicates
The known story of iodine in tunicates also appears to be complex. Tunicates apparently use the iodinated amino acids to make proteins, like the gorgonia, but they also make some very complicated organoiodine compounds.47-49 Since some of these molecules are vertebrate hormones (e.g., thyroxine) it has been suggested that tunicates use them in this fashion, while most lower invertebrates do not.48 Others of these iodoorganics are even more complex, such as lukianol B, a cytotoxic compound that might be made as a deterrent to predators.47 Like all of the species discussed above, tunicates have also been shown to take up iodine as dissolved inorganic iodine from the water column.49
Who Uses Iodine: Sponges
Sponges are also known to produce a wide variety of potentially toxic organoiodine compounds.50 One sponge of the genus Geodia is known to make cyclic peptides (geodiamolides A and B) that contain iodine, and these toxic compounds are expected to exert a protective effect of some type.51
Overall, the sponges do incorporate large amounts of iodine An analysis of 12 species of sponges from the Andaman Sea showed iodine contents that range from 0.001 to 0.085% iodine.52 I have not, however, seen any studies about where sponges get iodine, whether from inorganic iodine in the water column, or from ingested food.
Who Uses Iodine: Worms
The marine worm Glossobalanus minutus has been shown to take up iodine from the water column, but it is not clear what it does with it as no iodinated proteins were detected.53 A different marine worm, Lineus rubber, accumulates iodide from the water column and stores it mostly as iodide, with a smaller amount of thyronine (an organoiodine hormone).54, 55
Who Uses Iodine: Shrimp
As one moves up the food chain to more sophisticated organisms, data on their ability to take up iodine from the water column becomes very sparse. Shrimp are known to have a need for iodine to grow, but that is only known in terms of a dietary requirement. The shrimp, Penaeus chinensis O’sbeck, for example, grows optimally when the diet contains 0.003% iodine.56
Shrimp apparently incorporate substantial amounts of iodoorganics into their bodies. The shrimp Pandalus borealis, for example, incorporates between 0.04 and 2 ppm iodine as iodoorganic compounds depending on the particular body tissues examined. Their roe were somewhat higher, up to 4 ppm iodine as iodoorganic compounds.57 Shrimp shells and other parts can contain up to 17 ppm by dry weight iodine, the majority of which is iodoorganic compounds58, but the values are still far lower than for other inverts like macroalgae, sponges, or gorgonia.
Still, the amount contained says nothing about whether iodine is an important requirement. I could find no scientific studies that showed that shrimp need iodine from the water column, but neither could I find any that demonstrates that they do not.
Who Uses Iodine: Soft Corals Such As Xenia
Surely, you say, there must be studies showing that Xenia and other soft corals need iodine from the water column? Well, I could find none. There may be studies that I could not find, and regardless of whether there are studies, iodine in the water column may or may not have a significant impact on these organisms. Nevertheless, there is no published basis (that I could find) for many of the claims about iodine.
There are studies that show that Xenia does contain substantial iodine, and it is likely that it got it from the water column, but what good, if any, that iodine serves is unknown. In a recent publication, Ron Shimek showed that a wild specimen of Xenia sp. contained 350 ppm iodine on a wet basis and a captive specimen showed 270 ppm on a wet basis and 1350 ppm on a dry basis.29 Those values are as high as some of the macroalgae, and lend some support to the idea that Xenia accumulate iodine (and presumably have a use for it at such high accumulations).
Of course, accumulating iodine from the tank somehow, and showing that supplemental inorganic iodine is beneficial are very different. I am in the planning stages of running experiments on the possible benefits of iodine supplementation to certain soft corals, but the technical challenges are significant (much more so than similar tests on macroalgae), and I’m not certain that they will be successful.
Measurement of Iodine: Test Kits
There are many ways to measure iodine in seawater. Even confining the list to those that most aquarists are likely to encounter, there are still at least three fundamentally different ways, and those three all detect different subsets of the total iodine species in water. Unfortunately, those differences make comparisons of the different methods almost impossible, and also complicate the interpretation of the data from any single one of them. Of the commercially available kits, the Seachem and Salifert iodine kits are the most popular in the US. Unfortunately, the use and interpretation of these kits is tedious and complicated. I’d like to hold off on detailed comments about how well they work until I’ve had additional time to study them, but so far my experience has been rather less successful that with simpler kits (alkalinity, calcium, etc.).
The Seachem iodine kit detects only iodide (I–) and molecular iodine (I2). It is unlikely that there will be much I2 in an aquarium as it quickly breaks down into other products59 [to be described in a later article that will include a discussion on using products that contain I2, such as Lugols solution]. A potential drawback to this kit is that it doesn’t detect iodate. So if your tank water matches normal seawater in quantity and speciation of iodine, then it will look artificially low (say, about 0.01 or 0.02 ppm). I would also not advise using this kit if you are adding an iodine supplement that contains considerable amounts of iodate, molecular iodine (that may break down into products that include iodate and iodide)59, or organic iodine forms. There is also the concern that supplemental iodide, and those forms of iodine coming in with marine foods, may end up partially as iodate. Consequently, this kit may substantially underestimate the total amount of iodine present. I fell into that trap years ago in dosing iodate to my tank, and a significant concentration built up before I specifically tested for it.
The Salifert iodine test kit detects iodide, iodate, and molecular iodine. Salifert is about to come out with a new iodine kit (using a yellow color instead of a pink color). Assuming that it functions properly, it would be a good choice for anyone dosing iodide, iodate, or molecular iodine, though it is a long test involving quite a few steps. It will not detect many organic iodine forms, and people dosing such compounds should beware of overdosing.
Measurement of Iodine: ICP
Another way of detecting iodine that aquarists sometimes encounter is via Inductively Coupled Plasma (ICP). In this machine intensive method, a water sample is sucked into a high temperature plasma. Iodine in all forms is vaporized and detected equally. In its most sensitive application, the plasma stream is sent into a mass spectrometer which counts the individual ions with a particular mass. This technique is referred to as ICP-MS. Data of that type has been reported to aquarists,60 with aquarium results ranging from 0.03 to 1.2 ppm iodine.
ICP-MS it is not the most common form of ICP used in routine analysis of water samples, however. The most standard way to detect what elements are present is with optical emission spectroscopy. In the high temperature plasma, the atoms emit light, and the wavelengths (colors) emitted are specific for each element. In an ICP using optical emission detection (referred to as ICP-OES), the detector is set to the wavelength(s) emitted by iodine, and the amount emitted at that wavelength is recorded and converted back to a concentration that must have been present in the water.
The wavelengths used for iodine detection in a standard ICP are all in the ultraviolet, with 178.215 nm being the normal first choice for iodine because of the strength of the emission at that wavelength. According Varian (the manufacturer of the ICP that I used to test my samples), the detection limit at that wavelength is about 0.6 ppm (far above natural seawater levels). This wavelength also has potential interference from phosphorus that emits near this wavelength (178.222 nm). The second choice line at 182.976 nm has a detection limit of 4.3 ppm. (again, according to Varian). So one can see that ICP using optical emission detection is not typically a good choice for measuring low levels of iodine (as in natural levels around 0.06 ppm), but can work for substantially elevated levels (greater than 0.6 ppm iodine).
I used ICP with optical emission detection to analyze unfiltered water from my tank (I have routinely dosed iodide to the tank, but had stopped about 3 weeks before taking this sample). A friend of mine who runs our analytical lab tested several samples for me with the sole purpose of searching for iodine. We were not able to detect any iodine in the tank water sample, nor were we able to detect any iodine in that same sample spiked with an additional 0.33 ppm iodide [For those interested, we used a Varian Vista MPX ICP-OES, and used pure deionized water as the baseline. We looked at both wavelengths described above, and could detect no iodine above the background noise present in the deionized water].
In Ron Shimek’s analysis of unfiltered water samples from aquaria, he used ICP-OES.61 He found that 38% of samples had no iodine detected above his stated background of 0.1 ppm iodine, but other samples had reported values up to 2.1 ppm iodine. These results are indicative of total iodine (iodide + iodate + organic iodine). The samples in that study with high iodine may have had elevated inorganic iodine forms (perhaps especially iodate which may deplete les rapidly from aquaria), or may have had substantial organic forms that would not be detected with test kits. It is also possible that some of those elevated values may represent interference in the test method by phosphorus, which was reported as strongly correlated with the reported iodine levels.62 In any case, one should be aware of the potential for overdosing of supplements, and even of iodine-containing foods. Using a test kit may help ensure that you do not develop this problem.
Sources Of Iodine In An Marine Aquarium
Other than iodine supplements, the significant sources of iodine in marine aquaria are likely to be almost exclusively in the foods given. Even though starting salt mixes contain some iodine, that source likely disappears very rapidly (it has been known for a long time that iodide depletes rapidly, and next month’s article will show some rates). So only ongoing additions of iodine are likely to have a significant long term impact. Most calcium and alkalinity supplements are expected to have little in the way of iodine in them, but I’ve not seen any quantitative analyses of them for iodine.
Of the foods provided to most marine aquaria, algae in fish food is likely to be a big source. Gracilaria sp. macroalgae are often added to tanks, both in commercial prepared foods (e.g., some of the Formula foods from Ocean Nutrition, which also includes additional inorganic iodide), and as an individual macroalgae for fish (e.g., Tang Heaven). In one study, Gracilaria sp. was found to contain 3654 ppm iodine by dry weight.27 So the daily input to a tank adding 3 grams (dry weight) of Gracilaria sp. per day is about 11 mg of iodine. That is enough to bring a 50-gallon tank from zero to natural levels (0.06 ppm iodine) EVERY DAY. In the end, I think that we should be very happy that iodine is depleted so rapidly, because if it were not, we’d probably rapidly drive up iodine levels in many tanks.
In a recent study of foods going into aquaria, Ron Shimek estimated that the average tank is his study added 27% of the total natural level of iodine every day, consistent with the above analysis.63 Yet many of the tanks (37%) showed no detectable iodine (by ICP which he stated has a 0.1 ppm level of detection) .61 Clearly, the additions are substantial.
What About Dosing?
It makes much more sense to give dosing recommendations after presenting iodine depletion rates and showing what impact, if any, supplementation has on some common organisms, so I will hold off on that all-essential discussion until next month. Nevertheless, there is one important piece of advice that I can give now.
I would strongly advise people to not try to maintain 0.06 ppm iodine using supplementation and a test kit.
Why you ask? Isn’t that what we do for most other chemicals of interest? Well, if we could easily and accurately determine the concentrations of the different forms of iodine in aquaria, then I would recommend doing just that. However, if you combine the complexities of having multiple iodine species present with the uncertainty of having hobbyists use very complex test kits that may readily yield incorrect or difficult to interpret results, the stage is set for people driving their total iodine to levels far from what they actually intended. I do not know what levels of iodide or iodate become apparently problematic to real aquaria, but the risk of overdosing in this fashion is not insignificant. Both of the ICP studies reported above found some tanks with substantially elevated total iodine levels. I have no way of knowing how those levels got there, but overdosing of a supplement is one likely way.
Why Such Different Reported Effects Of Iodine Supplementation?
There is a huge disparity among aquarists about the effects that they have seen in their tanks with and without dosing iodine. Some say they very clearly see certain effects when dosing iodine (an effect that often stops when they stop dosing), and other aquarists report not seeing those same effects. Often this is used as evidence that iodine supplementation cannot be having the effects observed in one tank if it does not have the same effect in another tank. Without claiming that iodine has any particular effect, I contend that such logic is flawed for the reason given below:
Since iodine appears to be fluxing massively through aquaria relative to the natural levels in seawater (close to 100% turnover every few days as noted above), it would not be surprising if, in the absence of supplementation, some tanks had much higher levels of iodine than others. While some tanks may stay pegged on the low side of 0.06 ppm without supplementation, it may be critical whether these are holding steady at 0.02 ppm, 0.001 ppm, or 0.00001 ppm. Few aquarists can detect and quantify these lower levels. Other tanks may have total iodine levels above 0.06 ppm due to feeding of high iodine foods. And even within the same total iodine level, different tanks, with or without supplementation, are going to have different relative amounts of iodide, iodate, and organic iodine. So the differences that people report in their tank’s response to iodine addition may be the result of:
- The fact that hobbyists are not especially scientific observers of their tanks, and they may be mistaken about the observed effects (or not) of supplementation.
- The natural amounts of the various species of iodine present in marine aquaria without supplementation vary considerably, and so the different observations reported may reflect different starting points. That is, some tanks may have enough iodine naturally present that additional iodine has no effect, while others may start so low that added iodine pushes the iodine level up to the point where certain new effects can be observed.
Do I know which effect is real? Or is it both? No, I do not know. But aquarists should be aware of these issues when discussing the effects of iodine supplementation. In next month’s article, I present some results of iodine supplementation on macroalgae growth. Regardless of the outcome, these experiments have little bearing on how iodine supplements impact other organisms, from shrimp to Xenia sp., but it does make a start at understanding what effects one might realistically expect from iodine supplementation.
Happy Reefing!
References
- The Halogens — Part III: Iodine. By Craig Bingman. Aquarium Frontiers. December 1997. http://www.animalnetwork.com/fish2/aqfm/1997/dec/bio/default.asp
- Iodide and iodate concentrations in eastern Australian subtropical waters, with iodide by ion chromatography. McTaggart, A. R.; Butler, E. C. V.; Haddad, P. R.; Middleton, J. H. Analytical Chemistry Department, University of New South Wales, Kensington, Sydney, NSW, Australia. Marine Chemistry (1994), 47(2), 159-72.
- Iodine speciation: A potential indicator to evaluate new production versus regenerated production. Tian, R. C.; Marty, J. C.; Nicolas, E.; Chiaverini, J.; Ruiz-Pino, D.; Pizay, M. D. Laboratoire de Physique et Chimie Marines, Observatoire Oceanologique, Villefranche-sur-Mer, Fr. Deep-Sea Research, Part I: Oceanographic Research Papers (1996), 43(5), 723-738.
- Dissolved organic iodine in marine waters: Determination, occurrence and analytical implications. Wong, George T. F.; Cheng, Xian-Hao. Department of Oceanography, Old Dominion University, Norfolk, VA, USA. Marine Chemistry (1998), 59(3-4), 271-281.
- The transformation of iodate to iodide in marine phytoplankton cultures. Wong, George T. F.; Piumsomboon, Ajcharaporn U.; Dunstan, William M. Department of Ocean, Earth and Atmospheric Sciences, Old Dominion University, Norfolk, VA, USA. Marine Ecology: Progress Series (2002), 237 27-39.
- Coupling iodine speciation to primary, regenerated or “new” production: a re-evaluation. Wong, George T. F. Department of Ocean, Earth and Atmospheric Sciences, Old Dominion University, Norfolk, VA, USA. Deep- Sea Res., Part I (2001), 48(6), 1459-1476.
- Reduction of iodate in seawater during Arabian Sea shipboard incubations and in laboratory cultures of the marine bacterium Shewanella putrefaciens strain MR-4. Farrenkopf, Anna M.; Dollhopf, Michael E.; Chadhain, Sinead Ni; Luther, , George W., III; Nealson, Kenneth H. University of Delaware, College of Marine Studies, 700 Pilottown Road, Lewes, DE, USA. Mar. Chem. (1997), 57(3-4), 347-354.
- Absorption and elimination of chemical forms of iodine-131 by marine macrophytes. Svetasheva, S. K. Inst. Biol. Yuzhn. Morei, Sevastopol, USSR. Gidrobiologicheskii Zhurnal (1984), 20(4), 94-6.
- Chemical forms of radioactive iodine in seawater and its effects upon marine organisms. Hirano, Shigeki; Ishii, Toshiaki; Nakamura, Ryoichi; Matsuba, Mitsue; Koyanagi, Taku. Natl. Inst. Radiol. Sci., Nakaminato, Japan. Radioisotopes (1983), 32(7), 319-22.
- Effect of stable iodine upon the concentration of radioactive iodine by marine organisms. Hirano, Shigeki; Matsuba, Mitsue; Koyanagi, Taku. Div. Mar. Radioecol., Natl. Inst. Radiol. Sci., Nakaminato, Japan. Radioisotopes (1983), 32(8), 353-8.
- Model experiments on the diurnal cycling of iodine in seawater. Brandao, Ana Claudia M.; Wagener, Angela de Luca; Wagener, Klaus. Dep. Quimica, Pontificia Univ. Catolica, Rio de Janeiro, Brazil. Mar. Chem. (1994), 46(1-2), 25-31.
- Antibacterial properties of starch iodide and of its components. Mokhnach, V. O.; Litvinov, M. A.; Borisov, L. B.; Matyko, N. A.; Smirnova- Ikonnikova, M. I. V. L. Komarov Botan. Inst., Leningrad, Mikrobiologiya (1960), 29 451-4.
- Preservation of potassium iodide eye drops with thiomersal. Stock, Wilfried; Klaus, Uwe. Zentrallab. Deutscher Apotheker, Eschborn, Fed. Rep. Ger. Pharmazeutische Zeitung (1982), 127(13), 733-6.
- Control of the amount of free molecular iodine in iodine germicides. Hickey, John; Panicucci, Rick; Duan, Yongjun; Dinehart, Kirk; Murphy, Joe; Kessler, Jack; Gottardi, Waldemar. Symbollon Corporation, Framingham, MA, USA. Journal of Pharmacy and Pharmacology (1997), 49(12), 1195-1199.
- Toxicity of iodine, iodide, and iodate to Daphnia magna and rainbow trout (Oncorhynchus mykiss). Laverock, M. J.; Stephenson, M.; Macdonald, C. R. AECL Res., Whiteshell Laboratories, Pinawa, MB, Can. Archives of Environmental Contamination and Toxicology (1995), 29(3), 344-50.
- Effect of potassium iodide and bromide on rainbow trout. Goreva, V. A. Saratov. Otd., GosNIORKh, Saratovo, USSR. Gidrobiologicheskii Zhurnal (1984), 20(5), 50-4.
- The transformation of iodate to iodide in marine phytoplankton cultures. Wong, George T. F.; Piumsomboon, Ajcharaporn U.; Dunstan, William M. Department of Ocean, Earth and Atmospheric Sciences, Old Dominion University, Norfolk, VA, USA. Marine Ecology: Progress Series (2002), 237 27-39.
- Effects of iodide and iodate ions on marine phytoplankton. Fuse, Hiroyuki; Takimura, Osamu; Yamaoka, Yukiho. Gov. Ind. Res. Inst., Kure, Japan. Editor(s): Okaichi, Tomotoshi; Anderson, Donald M.; Nemoto, Takahisa. Red Tides: Biol., Environ. Sci., Toxicol., Proc. Int. Symp., 1st (1989), Meeting Date 1987, 229-32. Publisher: Elsevier, New York, N. Y
- Biochemistry of iodine. I. The distribution of iodine in plant and animal tissues. Cameron, A. T. Univ. Manitoba, J. Biol. Chem. (1914), 18 335-80.
- Seasonal variation, bioaccumulation and prevention of loss of iodine in seaweeds. Mairh, O. P.; Ramavat, B. K.; Tewari, A.; Oza, R. M.; Joshi, H. V. Cent. Salt Mar. Chem. Res. Inst., Bhavnagar, India. Phytochemistry (1989), 28(12), 3307-10.
- Biological aspects of iodine behavior during interaction of algae with sea water. Kravtsova, Yu. Yu.; Saenko, G. N. USSR. Editor(s): Krasnov, E. V. Vzaimodeistvie Vodoi Zhivym Veshchestvom, Tr. Mezhdunar. Simp. (1979), Meeting Date 1975, 1 146-52.
- Influence of iodine on growth and development of the brown alga Ectocarpus siliculosus in axenic cultures. Woolery, Marti L.; Lewin, Ralph A. Scripps Inst. Oceanogr., Univ. California, La Jolla, CA, USA. Phycologia (1973), 12(3-4), 131-8.
- Environmental control of gametogenesis in Laminaria saccharina. III. Effects of different iodine concentrations and chloride and iodide ratios. Hsiao, Stephen I.; Druehl, Louis D. Dep. Biol. Sci., Simon Fraser Univ., Burnaby, BC, Can. Canadian Journal of Botany (1973), 51(5), 989-97.
- Influence of iodine and bromine on growth of some red algae in axenic culture. Fries, Lisbeth. Univ. Uppsala, Swed. Physiol. Plantarum (1966), 19(3), 800-8.
- Effects of iodine on the growth of the fronds of Asparagopsis armata (Rhodophyceae, Bonnemaisoniales) in culture from spear bearing branches. Codomier, Louis; Segot, Madeleine; Combaut, Georges. Groupe Rech. Biol. Chim. Veg. Mar., Cent. Univ. Perpignan, Perpignan, Fr. Giornale Botanico Italiano (1979), 113(5-6), 387-93.
- Concentration of iodine and bromine by plants in the seas of Japan and Okhotsk. Saenko, G. N.; Kravtsova, I. I.; Ivanenko, V. V.; Sheludko, S. I. Far East Sci. Cent., Inst. Chem., Vladivostok, USSR. Marine Biology (Berlin, Germany) (1978), 47(3), 243-50.
- Distribution of iodine in marine algae of Goa region. Solimabi; Das, B. Natl. Inst. Oceanogr., Goa, India. Indian Journal of Marine Sciences (1977), 6(2), 180-1.
- Instrumental neutron activation analysis of iodine levels in fourteen seaweed species from the coastal belt of Ghana. Serfor-Armah, Y.; Nyarko, B. J. B.; Carboo, D.; Osae, E. K.; Anim-Sampong, S.; Akaho, E. H. K. Chemistry Department, National Nuclear Research Institute, Ghana Atomic Energy Commission, Legon-Accra, Ghana. Journal of Radioanalytical and Nuclear Chemistry (2000), 245(2), 443-446.
- Down the Drain; Exports from Reef Aquaria by Ronald L. Shimek http://reefkeeping.com/issues/2002-12/rs/feature/index.htm
- Absorption and elimination of chemical forms of iodine-131 by marine macrophytes. Svetasheva, S. K. Inst. Biol. Yuzhn. Morei, Sevastopol, USSR. Gidrobiologicheskii Zhurnal (1984), 20(4), 94-6.
- Determination of chemical species of iodine in some seaweeds (I). Hou, Xiaolin; Chai, Chifang; Qian, Qinfang; Yan, Xiaojun; Fan, Xiao. Laboratory Nuclear Analysis Techniques, Academia Sinica, Institute High Energy Physics, Beijing, Peop. Rep. China. Science of the Total Environment (1997), 204(3), 215-221.
- Fate of iodides taken up by various marine algae and characterization of the iodinated amino acids in the hydrolysates. Andre, S. Lab. Biochim. Gen. Comp., Coll. France, Paris, Fr. Comptes Rendus des Seances de la Societe de Biologie et de Ses Filiales (1971), 165(12), 2293-8.
- Iodine-containing complexes of the Black Sea alga Phyllophora nervosa. Gazha, P. A.; Yunusov, T. S.; Shadrina, T. Yu.; Andrianov, A. M. Inst. Khim. Rastit. Veshchestv, Tashkent, USSR. Khimiya Prirodnykh Soedinenii (1983), (6), 772-6.
- Haloforms in the essential oil of the alga Asparagopsis taxiformis. Burreson, B. J.; Moore, Richard E.; Roller, Peter. Dep. Chem., Univ. Hawaii, Honolulu, HI, USA. Tetrahedron Letters (1975), (7), 473-6.
- New iodolactone from Laurencia majuscula. Su, Jing-Yu; Xu, Xiao-Hua; Zeng, Long-Mei; Wang, Chao-Jie. Dep. Chem., Zhongshan Univ., Canton, Peop. Rep. China. Gaodeng Xuexiao Huaxue Xuebao (1997), 18(8), 1333-1334.
- Marine macroalgae in polar regions as natural sources for volatile organohalogens. Laturnus, Frank. Department of Plant Biology and Biogeochemistry, Riso National Laboratory, Roskilde, Den. Environmental Science and Pollution Research International (2001), 8(2), 103-108.
- The involvement of hydrogen peroxide in the production of volatile halogenated compounds by Meristiella gelidium. Collen, Jonas; Ekdahl, Anja; Abrahamsson, Katarina; Pedersen, Marianne. Department of Physiological Botany, Uppsala University, Uppsala, Swed. Phytochemistry (1994), 36(5), 1197-1202.
- Production of halocarbons from seaweeds: an oxidative stress reaction? Pedersen, Marianne; Collen, J.; Abrahamsson, K.; Ekdahl, A. Department of Physiological Botany, Uppsala University, Uppsala, Swed. Scientia Marina (1996), 60(Supl. 1, Underwater Light and Algal Photobiology), 257-263.
- Release of Volatile Iodinated C1-C4 Hydrocarbons by Marine Macroalgae from Various Climate Zones. Giese, Bernd; Laturnus, Frank; Adams, Freddy C.; Wiencke, Christian. Department of Chemistry, University of Antwerp (UIA), Wilrijk, Belg. Environmental Science and Technology (1999), 33(14), 2432-2439.
- Biogenic volatile organoiodine and organobromine hydrocarbons in the Atlantic Ocean from 42°N to 72°S. Schall, C.; Heumann, K. G.; Kirst, G. O. Institute Inorganic Analytical Chemistry, Johannes Gutenberg- University, Mainz, Germany. Fresenius’ Journal of Analytical Chemistry (1997), 359(3), 298-305.
- Phytogenesis of halomethanes: A product of selection or a metabolic accident? Manley, Steven L. Department of Biological Sciences, California State University, Long Beach, CA, USA. Biogeochemistry (2002), 60(2), 163-180.
- 42 Iodine in gorgonian corals. Sugimoto, Koichi. J. Biol. Chem. (1928), 76 723-8.
- Chemical changes accompanying maturation of the connective tissue skeletons of gorgonian and antipatharian corals. Goldberg, W. M. Dep. Biol. Sci., Florida Int. Univ., Miami, FL, USA. Marine Biology (Berlin, Germany) (1978), 49(3), 203-10.
- Iodine content of the scleroproteins forming the horny stems of the Gorgonidae. Lafon, Marcelle; Mayol, Francoise. Compt. rend. soc. biol. (1948), 142 360-2.
- Chemical composition of the sclerotized black coral skeleton (Coelenterata: Antipatharia): a comparison of two species. Goldberg, Walter M.; Hopkins, Theodore L.; Holl, Susan M.; Schaefer, Jacob; Kramer, Karl J.; Morgan, Thomas D.; Kim, Kiho. Dep. Biol. Sci., Florida Int. Univ., Miami, FL, USA. Comparative Biochemistry and Physiology, Part B: Biochemistry & Molecular Biology (1994), 107b(4), 633-43.
- Thyroxine and vitamin D in the gorgonian Leptogorgia virgulata. Kingsley, R. J.; Corcoran, M. L.; Krider, K. L.; Kriechbaum, K. L. Department of Biology, University of Richmond, Richmond VA, USA. Comparative Biochemistry and Physiology, Part A: Molecular & Integrative Physiology (2001), 129A(4), 897-907.
- A complex pyrrolo-oxazinone and its iodo derivative isolated from a tunicate. Yoshida, Wesley Y.; Lee, Kit K.; Carroll, Anthony R.; Scheuer, Paul J. Dep. Chem., Univ. Hawaii, Manoa, Honolulu, HI, USA. Helvetica Chimica Acta (1992), 75(5), 1721-5.
- Presence and biosynthesis of thyroid hormones in a tunicate, Ciona intestinalis. Roche, Jean; Salvatore, Gaetano; Rametta, Giuseppe. Univ. Stazione Zoologica, Naples, Biochim. Biophys. Acta (1962), 63 154-65.
- Presence of thyroid hormone in a pelagic tunicate, Salpa maxima. Roche, Jean; Rametta, Giuseppe; Varrone, Stelio. Univ. Naples, Compt. Rend. Soc. Biol. (1962), 156 1964-8.
- Bromo- and iodo-containing alkaloids from marine microorganisms and sponges. Dembitsky V M Department of Pharmaceutical Chemistry and Natural Products, School of Pharmacy, Hebrew University of Jerusalem, Jerusalem, 91120 Israel. [email protected] BIOORGANICHESKAIA KHIMIIA (2002 May-Jun), 28(3), 196-208.
- Stereostructures of geodiamolides A and B, novel cyclodepsipeptides from the marine sponge Geodia sp. Chan, Wilfred R.; Tinto, Winston F.; Manchand, Percy S.; Todaro, Louis J. Dep. Chem., Univ. West Indies, St. Augustine, Trinidad/Tobago. Journal of Organic Chemistry (1987), 52(14), 3091-3.
- Bromine and iodine content in sponges and algae of the Andaman Sea. Solimabi, B. Das; Mittal, P. K.; Kamat, S. Y. Natl. Inst. Oceanogr., Goa, India. Indian Journal of Marine Sciences (1981), 10(3), 301-2.
- Fixation of iodine by an enteropneust (Glossobalanus minutus) and its significance. Roche, Jean; Rametta, Giuseppe; Salvatore, Gaetano. Univ. Naples, Compt. Rend. Soc. Biol. (1961), 155 1902-6.
- Iodine accumulation in a nemertine, Lineus ruber. Balfour, W. E.; Willmer, Edward N. Univ. Cambridge, Cambridge, UK. Journal of Experimental Biology (1967), 46(3), 551-6.
- Organic binding of iodide in nemertean mucus. Major, C. W.; Hanegan, J. L.; Anoli, Louis. Univ. of Maine, Orono, ME, USA. Comparative Biochemistry and Physiology (1969), 28(3), 1153-60.
- Requirements of shrimp, Penaeus chinensis O’sbeck for potassium, sodium,magnesium and iodine. Liu Fa Yi Li He Fang; Wang Hui Liang; Liang De Hai; Tian Yu Chuan Chinese Journal of Oceanology and Limnology. 1995; 13 (2) 141-146.
- Spatial distribution of extractable organohalogens in northern pink shrimp in the North Atlantic. Bottaro, Christina S.; Kiceniuk, Joe W.; Chatt, Amares. Trace Analysis Research Centre, Department of Chemistry, Dalhousie University, Halifax, NS, Can. Biological Trace Element Research (1999), 71-72 149-166.
- Iodine content of shrimp waste. Parker, Haywood, Jr.; Vilbrandt, Frank C. J. Am. Chem. Soc. (1931), 53 633-5.
- Disproportionation and reduction of molecular iodine added to seawater. Truesdale, Victor W.; Canosa-Mas, Carlos E.; Luther, George W. III. College of Marine Studies, University of Delaware, Lewes, DE 19958-1298, USA. Marine Chemistry (1995), 51(1), 55-60.
- The modern coral reef aquarium. Volume 1. Fossa. S. A. and Nilsen, A. J.
- It’s (in) the water. By Ronald Shimek: http://reefkeeping.com/issues/2002-02/rs/feature/index.htm
- It is still in the water. By Ronald Shimek: http://reefkeeping.com/issues/2002-03/rs/feature/index.htm
- What we put in the water. By Ronald Shimek: http://reefkeeping.com/issues/2002-04/rs/feature/index.htm

Extremely interesting article as I am about to start dosing iodine on my sps tank after my icp test found it very low,will be dosing at a lower rate than is required for six weeks and then test again,thank you for your help on this subject.